Was
Darwin
Wrong?
Was Darwin Wrong? |
Home | Intro | About | Feedback | Prev | Next |
|
 |
by Gert Korthof, 3 Oct 2006 (updated 22 Mar 2009: 34 )
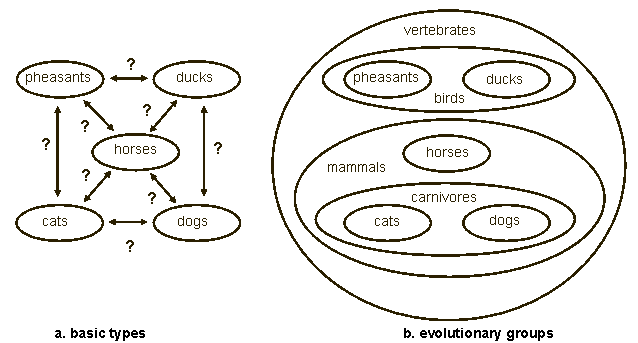
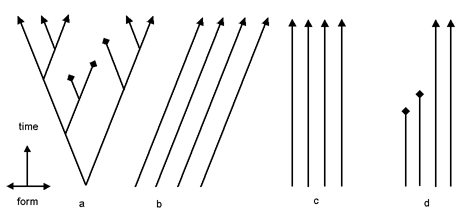
Is it possible to accept micro-evolution and reject macro-evolution? Is a compromise possible between creation and evolution? I will ignore the supernatural character of creation models, and argue that anything less than full common descent leads to an arbitrary fragmentation of the tree of life, leads to a logically inconsistent theory of descent, and conflicts with the evidence. The Swedish botanist, Carl Linnaeus, who invented the biological nomenclature that is still in use today, was a creationist. According to Linnaeus: "We count as many species as different forms were created in the beginning" (Mayr 1982, 258 14). Linnaeus's work and thinking were based on the concept of design by God. Species were fixed, and their systematic relationships reflected the divine plan. Despite his belief in the fixity of species, Linnaeus came to accept there are varieties within species. These varieties, he believed, resulted from changed conditions. In 1756, at the end of his life, he came to believe that the number of species within a genus might increase. This was not evolution as we know it today; Linnaeus thought that new species were produced by hybridization of existing species. Nevertheless the effect was that the fixity of species had become somewhat less than fixed (Bowler 1989, 67 3). The French naturalist, Georges-Louis Buffon, had defended the fixity of species early in his career, but in 1766, nearly a hundred years before the publication of Darwin's Origin of Species, he accepted the idea that closely related Linnaean species had diverged from a common ancestor. This view is close to what today is called microevolution. Buffon even went as far as to claim that families were created by God. The family possessed fixed characteristics and had no ancestors itself (Bowler 1989, 74 3). Today, philosopher Paul Nelson, who is part of the Intelligent Design movement, argues for the creation of basic types ("ground types") stemming from common ancestors (Nelson 2001:684 16). He illustrates basic types with a figure from Junker and Scherer (Fig. 1a, 32).
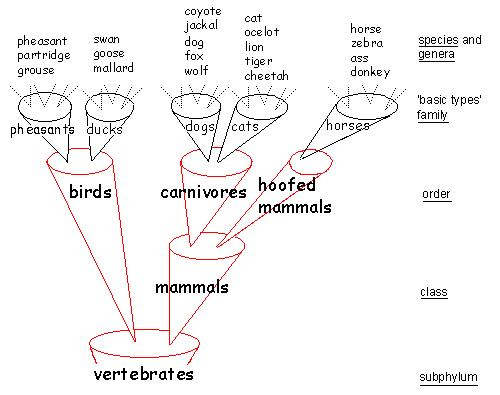
Figure 1b shows five animal groups: pheasants, ducks, dogs, cats, and horses. Each group is descended from a created common ancestor, which itself has no ancestor. Nelson contrasts a static-creation theory (creation of fixed species) with the dynamic-creation model he favors. Nelson criticizes Mark Ridley (1985 21) for displaying only the static fixed-species model and ignoring the modern dynamic-creation model (Fig. 2).
The four publications Nelson uses as evidence for his accusation, including the source of Figure 1a (Junker and Scherer 1988), postdate Ridley's 1985 book. This is an unfair criticism, because Ridley could not have known of publications appearing after his own book. Nelson is not the only creationist proposing this kind of creation model. Jonathan Sarfati (2000, 38, 39 26) has a similar model called "the true creationist orchard" (the picture of the model looks like an orchard). The model says that diversity has occurred within the original Genesis kinds. There are no names attached to his trees in his illustration, so no inference as to the taxonomic level of the trees can be drawn. He too contrasts his model with the alleged creationist "lawn." The fixed-species model claims that no new species have been produced since creation, but it allows extinction. This is a caricature of creationism, says Sarfati, because it implies that the Genesis kinds were the same as today's species. Sarfati claims especially that the fixed-species model is a straw man for creationism. Nelson also claims that it does not occur in creationist publications of the last decades. This statement implies that creationists adhered to the fixed-species model only a few decades ago. It would be interesting to know what insight or fact caused their conversion to the dynamic-creation model. The dynamic-creation model is similar to the model of the creationist Walter Remine (1993, 444 23). He, for example, places dogs, wolves, coyotes, jackals, and foxes in one systematic group: the Canidae-monobaramin. This matches Nelson's dogs group. Remine defines monobaramin as "a group containing only organisms related by common descent, sharing a common ancestor" (444). And subsequently Remine states the inevitable: "Directly created organisms have no ancestor, they are created by the direct action of a designer" (510).
Implications of the Dynamic-Creation ModelThe dynamic-creation model (DCM) uses the theological concept of creation as its foundation: "Here, the terminal species are members of basic types, stemming from common ancestors which were themselves created" (Nelson 2001:684 16). Therefore the DCM is basically theology. Nelson could have omitted the word "creation." He could have used an agnostic formula such as "the common ancestors of families are unknown and cannot be known to science," but he did not. Doing so would have destroyed the beating heart of creation theory. In the following I will ignore the theological foundation of the DCM, not because it is a minor detail, but to side-step accusations of atheism and avoid endless discussions about whether supernatural interventions are a legitimate part of natural science. I will first explore the biological implications of the DCM and then evaluate the model itself.
Evaluation of the Dynamic-Creation Model (DCM)MysteriesThe dynamic-creation model uses standard neo-Darwinian processes when convenient but also introduces mysteries and fatal inconsistencies. Let's first have a look at the orthodoxy. The mini-trees imply common descent, branching evolution, hierarchical taxonomic levels, origin of new species, natural selection and mutation. For example, the model explains similarities within basic types (similarities of dogs, wolves, foxes, and coyotes) by common descent. Additionally, the differences between dogs, wolves, foxes, and coyotes are explained by divergence of the organisms arising from the ancestral basic type. Both facts are reflected in the mini-trees. So far, so good. But then a huge difference from the standard Darwinian explanation arises: in the DCM, cats and dogs have an independent origin. Cats and dogs are completely unrelated groups. They are not related by common descent. That destroys the standard (Darwinian) explanation of their similarities.
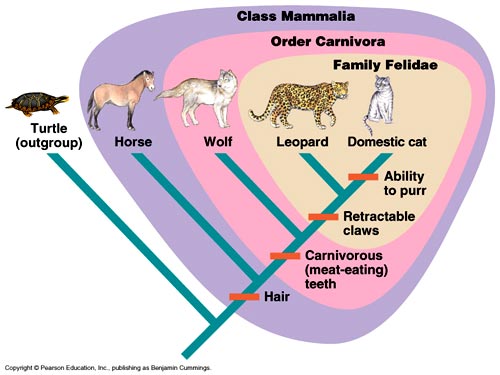
InconsistencyNow the logical inconsistency is easy to see. If the similarities and dissimilarities are a good reason for classifying individual organisms into the hierarchical categories, species, genera, and families, and for explaining that pattern with common descent, then why are those reasons not equally valid for higher categories such as orders, classes, phyla, and kingdoms? Why is common descent a good explanation up to the family level and a bad explanation at higher levels? Even horses and birds share vertebrate characteristics. This pattern of similarities is called the groups-within-groups pattern, or inclusive groups (Fig. 3b; Fig. 4.). To return to Nelson's mini-trees: the dogs are a group within the Carnivores group, the Carnivores are a group within Mammalia, and the Mammalia are a group within the Craniata, and the Craniata are a group within the Animalia. The logic of inclusive groups makes it impossible to see them as independent groups. Every taxonomic group (except the highest) is included in a higher-level group. There is no such a thing as an independent group.
Animal FarmAccording to the DCM, basic types have an independent origin. That means basic types are not connected by common descent. All basic types are equal in the sense that they are equally independent. To paraphrase George Orwell ("all animals are equal, but some animals are more equal than others"), all basic types are unequal, but some basic types are more unequal than others. Indeed, there are degrees of similarities. Although, for example, the three basic types, pheasants, ducks, and cats, have an independent origin, creationists cannot deny that pheasants and ducks are more equal than pheasants and cats. But why should some basic types look more similar than others, if there is not a close evolutionary relationship between them? And why should some basic types look less similar, if there is not a more-distant evolutionary relationship between them? Why expect any similarities above the basic-type level at all? Any pattern of similarities of basic types is possible. Independent origin is unable to predict a groups-within-groups pattern. Higher-level groups, such as birds, carnivores, mammals, reptiles, fishes, insects, and plants, are not expected or predicted at all by a theory of independent origin. In fact, relations between basic types are in principle unknowable (Fig. 3a), whereas evolutionary relations between groups become clearer when new information becomes available.
What can be concluded from the dynamic-creation model is that the unity of living things is a hoax and their diversity is a joke. Scherer (1998, 206 25) tries to give an empirical definition of the basic-type. The point, however, is that he does nothing to establish the similarities and degrees of similarity between basic types. Every biologist classifies pheasants and ducks in one group, Aves (birds), and explains their similarity by saying that pheasants and ducks have a more-recent ancestor than pheasants and cats. The explanation of the Linnaean hierarchical classification system collapses if we accept the independent origin of basic types.
Plan of CreationIn Linnaeus's time, the existence of the groups-within-groups pattern was "explained" as the plan of creation. It was, in reality, an unintelligible fact. Without evolution, nobody could hope for a better explanation. Maybe the need for an explanation was not even felt. But it is unacceptable to fall back to the mysteries of pre-Darwinian times when there is a good explanation available. It is not exactly progress in science to introduce mysteries, inconsistencies, and the destruction of the explanation of all the taxonomic categories above the family level. Darwin offered an elegant explanation for the groups-within-groups pattern: common descent. The creation model leaves unexplained or completely mysterious all those similarities that common descent elegantly and consistently explains. Even allowing for creation as a scientific explanation still leaves the pattern of similarities unexplained. The assumption of created ancestors does not lead to specific expectations about the pattern of life on earth such as the groups-within-groups pattern. How could it? "Darwin, after all, banished speculation about the "unknown plan of creation" from science" (Johnson 1993, 70 10)).
Reinventing Common DescentScherer (1998 25) tried to express biological relationships between species into a new systematic category: basic types. This category will not help to classify organisms belonging to different basic types. To capture the relationships between pheasants, ducks, and all the other birds, one needs a basic type called birds, which is the ancestor of all the different basic types of birds. The same holds for all the mammals and all the animals. One needs basic types called "mammals" and "animals." To capture the relationships between all forms of life on earth, one needs a basic type called "life." And that amounts to reinventing common descent. The implication is that Nelson's and Scherer's basic types are neither "basic" nor "types." Mutation and natural selection can go beyond the confines of basic types, unaware of the boundaries of basic types.
Darwin's InventionNelson's criticism of Ridley's illustration of different histories of species on earth (Ridley, 1985) is odd for many reasons. The dynamic-creation model contains mini-trees. But the idea to use trees to represent the relations between species is stolen from Darwin and certainly not invented by creationists:"Darwin, curiously, was the first author to postulate that all organisms have descended from common ancestors by a continuous process of branching. . . .. A continuing multiplication of species could account for the total diversity of organic life" (Mayr 1982, 507 14).Additionally, the trees in Ridley's illustration have diverging branches, while those in Nelson's illustration are vertical. That means that species are static. The dynamic-creation model is a distortion of common descent. One cannot even say that the DCM is halfway toward common descent. Nelson and other intelligent-design theorists are blind to the power and purpose of common descent. Common descent does not explain families; it explains life. To limit the scope of common descent to families is like driving a plane on the road and ignoring that a plane is meant to fly!
New Evidence for Common Descent of "Basic Types"So far I have focussed on logic and explanatory power. These can be understood without detailed knowledge of genetics and biochemistry. Indeed, Darwin knew nothing about genetics and biochemistry, but he understood the logic of evolutionary theory. In 1900, Mendelian genetics was born. Fifty years later, James Watson and Francis Crick published the structure of DNA. Another 50 years later, the complete sequence of the human genome was published. The last event signified the transformation of genetics (the study of individual genes) into genomics (the study of the genome, or the whole gene set of a species). When scientists started comparing whole genomes of species, startling new evidence for the similarity and common descent of species began to emerge. The DNA sequences of human and mouse, for example, revealed that not only genes, but also whole segments of chromosomes of mouse and man are identical. A chromosome segment of roughly 90.5 million DNA bases on human chromosome 4 is similar to mouse chromosome 5. Almost all human genes on chromosome 17 are found on mouse chromosome 11, and human chromosome 20 corresponds entirely to the bottom segment of mouse chromosome 2. A graphical and interactive representation of all the cases of synteny (correspondence of chromosome segments of two different species) of mouse and man can be found at the Web site of the Sanger Institute (2002). The maps are based on (Gregory et al 2002 9). These special similarities mean that hundreds to thousands of genes are found in the same order in both mouse and human. This is impressive evidence for their common descent. The distribution of genes over chromosomes cannot be explained by biochemical or biological necessity. With the exception of the Y-chromosome and to a lesser degree the X-chromosome, no chromosome is dedicated to a special biological function such as digestion, respiration, locomotion, reproduction, or perception. The genes that control those biological functions are distributed over 20 pairs of chromosomes in the mouse and 23 pairs in the human. Consequently, the correspondences between different species cannot be explained by necessity or chance. Historical contingency is the dominant factor that produced the size, shape, composition, and number of chromosomes. Despite the many rearrangements of chromosomes since the human-mouse split, both humans and mice inherited 195 intact, conserved segments from their common ancestor. In the creation model, man and mouse belong to different basic types. The correspondence of their chromosome segments shows, however, that mouse and man are not basic types but, rather, derived types. Similar results are found for cats, seals, cows, horses, and rats. Indeed, while I was working on the final revision of this chapter, Science magazine (13 June 2003) published a special issue devoted to the tree of life. Among the many enlightening topics was the visualization of a tree of life incorporating no less than 3000 species (Pennisi, 2003 19). Because of the immense amount of information packed into the tree, the editor noted, not without humor, "best viewed when enlarged to a 1.5 meter diameter." The latest stunning confirmation of common descent comes from the Human Genome project. The director of the project Francis Collins gave a splendid popular account of those findings (31).Since the Human Genome project the pace of phylogenetic discovery has accelerated to the point where nearly complete phylogenetic trees can be constructed for well-studied clades, such as mammals. Sequence data have been collected from nearly 182,000 eukaryotic species in the GenBank database. It is a matter of adding more data to construct the final high-resolution Tree of Life (33).
Michael BeheNot all intelligent-design advocates are like Nelson. Michael Behe (1996, 176 1) claims to accept the common descent of all life:"I believe the evidence strongly supports common descent."He repeated this statement in a later publication (Behe 2001, 697 2): ". . . since I dispute the mechanism of natural selection, not common descent."Behe's position is puzzling. Behe does not say why he accepts common descent. The two quotes are nearly all he has to say about common descent. I think Behe does not realize the consequences of his statement. Common descent of life means that all life on Earth is physically, historically, and genetically connected. Common descent of life means that life is one unbroken chain of ancestors and descendants. Common descent of life means that every organism inherited all its genes from the previous generation (with slight modifications). And that includes irreducibly complex systems. Every supernatural intervention is a violation of common descent, because it means that a new irreducibly complex system in the first individual showing it was not inherited from its parents. It would be unjustified to say, "I inherited all my chromosomes from my parents, except an irreducibly complex system on chromosome X, which has a supernatural origin." Equally Behe cannot claim that common descent is true except for the cases where irreducible complex systems show up. Common descent does not allow for that kind of exception, because it would imply a violation of the laws of genetics. Genetics is the most exact and well-established discipline within biology. Hundreds of thousands of genetic experiments have been done since the birth of classic Mendelian genetics (1900) and the birth of molecular genetics (1953). The sudden emergence of an irreducibly complex system has never been observed, whereas the kinds of mutations necessary for evolution have routinely been observed.
Phillip JohnsonPhillip Johnson is one of the leaders of the intelligent-design movement. His opinion about common descent is most clearly stated in his well-known Darwin on Trial:"Because their [creationists'] doctrine has always been that God created basic kinds, or types, which subsequently diversified. The most famous example of creationist microevolution involves the descendants of Adam and Eve, who have diversified from a common ancestral pair to create all the diverse races of the human species" (Johnson 1993, 68 10).This is an intriguing passage for several reasons. The passage suggests that humans are a "basic kind" and are created as such. By switching from "basic kind" to microevolution, Johnson avoids explicitly stating that humans are a basic kind and thus created. However, the passage strongly suggests that humans are a "basic kind." But if they are, then Johnson's idea of "basic kinds" is more restricted than Nelson's dynamic-creation model. Johnson's example of "basic kind" is at the species level and implicitly affirms that the meaning of "microevolution" is "change within a species." If this is Johnson's view, then by implication all species are created, and microevolution is allowed to create only minor modifications within species. Whatever the definition of basic kinds (be it on the species or family level), microevolution by definition produces no new species. Any creation model that ends up with more species than the number of species it started with needs a natural mechanism to produce new species. And creating new species is macroevolution according to the textbooks. But it is very difficult for a creationist to admit that he or she accepts macroevolution. From another passage we see that Johnson would love to believe in the special creation of humans: "We observe directly that apples fall when dropped, but we do not observe a common ancestor for modern apes and humans. What we do observe is that apes and humans are physically and biochemically more like each other than they are like rabbits, snakes, or trees. The ape-like common ancestor is a hypothesis in a theory, which purports to explain how these greater and lesser similarities came about. The theory is plausible, especially to a philosophical materialist, but it may nonetheless be false. The true explanation for natural relationships may be something much more mysterious (Johnson 1993, 67 10).Again an intriguing passage. Now Johnson states that the hypothesis that apes and humans share a common ancestor is plausible but may be false. He fails to make clear whether he accepts or rejects common descent and why. To state that a scientific theory may be false is nothing new, because all scientific theories may be false. Certainly the idea that the true explanation for relationships may be mysterious is not a solid reason to reject common descent. With humor Johnson remarks that "descent with modification could be a testable scientific hypothesis"(66). But suggesting a mysterious cause for natural relationships is not "a testable scientific hypothesis." I fail to see what philosophical materialism has got to do with it. Johnson does not propose a nonmaterialist explanation. Johnson evidently does not like the hypothesis of common descent, but is unable to give good reasons to reject it, and fails to present an alternative. This is science by personal preference.
William DembskiWilliam Dembski, the mathematician of the intelligent-design movement, published his main works after Johnson and Behe. However, he is less specific about common descent than creationists like Nelson and more ambivalent about the correctness of common descent than Michael Behe. The evolutionary biologist, H. Allen Orr (2002 18), stated that the intelligent-design movement usually admits that people, pigs, and petunias are related by common descent. However its leading theorist, William Dembski, does not unconditionally accept common descent (Dembski 2002:314, 315 6). He ignores Nelson's dynamic-creation model. He fails to say what, for example, the similarity of apes and humans means.Darwinism comprises a historical claim (common descent) and a naturalistic mechanism (natural selection operating on random variations), with the latter being used to justify the former. According to intelligent design, the Darwinian mechanism cannot bear the weight of common descent. Intelligent design therefore throws common descent into question but at the same time leaves open as a very live possibility that common descent is the case, albeit for reasons other than the Darwinian mechanism (Dembski 2002:315).Dembski is right in distinguishing between common descent of all life and the mechanism of evolution, but he is wrong about the relation between the two. Yes, both are part of Darwinism, but he is not right to suggest that natural selection and random variation are the justification for common descent. Darwin would have adopted his theory of common descent on the basis of classification alone. Common descent is inferred from data that are independent of the mechanism of evolution. Common descent does not necessarily imply anything about natural selection, neutralism, gradualism, punctuated equilibrium, or the fossil record. Dembski is determined to undermine the mechanism of evolution. He hopes to undermine common descent as a logical consequence of the destruction of the mechanism of evolution. Significantly, 150 years after Darwin, Dembski still has nothing better to say than a cryptic "time leaves open as a very live possibility that common descent is the case, albeit for reasons other than the Darwinian mechanism."This vague remark is very similar to Johnson's mysticism: "some process altogether beyond the ken of our science" (Johnson, 155). Remarkably, Johnson stated earlier in his book that "speculation is no substitute for scientific evidence" and that "Darwin, after all, banished speculation about the 'unknown plan of creation' from science" (70). It is too late for mysticism 150 years after Darwin. Biologists have something better: it is called common descent. Both Johnson and Dembski forget that Charles Darwin did not know about genetic mutations, Darwin did not know Mendelian genetics (rediscovered in 1900), and he did not know molecular genetics (born in 1953). The point is that Darwin had sufficient reason to explain patterns of similarities and dissimilarities in the organic world even without knowledge of genetics. The success of Darwin's explanation did not even depend on the specifics of his theory of heredity, which turned out to be wrong. Now that we know that the genetic language of all life (how genes are translated into proteins) is not only similar but also virtually identical in all organisms, we have a magnificent confirmation of common descent. Darwin could not have foreseen that common descent would receive such dramatic underpinnings. The specific genetic code that all living organisms use to translate genes into proteins could have been dramatically different; no chemical laws that render the current genetic code necessary have been discovered. Each created basic type could have a different genetic code without any physiological or ecological problems (Korthof 2001). Dogs and cats could have different genetic codes. Humans and apes could have different genetic codes. Yet, they do not. Common descent would be best refuted if the most closely related organisms had the most dissimilar genetic codes (theoretically, genetic codes can differ in gradual ways). But all species have the same genetic code. The rare and small variations of the genetic code are superimposed on common descent and follow the pattern of descent with modification. Phylogenetic trees can be constructed for those variants. The genetic code, which translates genes into proteins, is stored in DNA and subject to mutation. Variant genetic codes have very restricted effects on the organism. Nearly all possible variant genetic codes are destructive for the organism and are subject to very strong selection pressure. That explains why variations are rare. Far from being an argument against common descent, as some creationists argue, they give clues for the origin of the genetic code. Furthermore, Darwin did not and could not construct the theory of common descent to explain the universality of the genetic code. The universality of the genetic code was discovered more than one hundred years after the publication of the Origin of Species. Common descent thus successfully explains a completely new fact about life on earth. All those similarities will not go away, whatever Dembski's claims about the inadequacy of the mechanism of evolution. In fact, it is extremely hard to come up with a complete and systematic alternative to common descent. Attempts to formulate a naturalistic alternative to common descent have resulted in severe problems and absurdities. For example, the Senapathy / Schwabe hypothesis of independent origin has even greater problems in explaining the properties of life. See for the full story about Dembski's position on Common Descent (2007) this Pandas Thumb post.
Is There an Alternative to Common Descent?Is there an alternative to common descent, and can there be partial common descent? The dynamic-creation model (DCM), with its created types and mini-trees, breaks the living world into arbitrary fragments, whereas common descent unifies all life. In fact, common descent unifies all disciplines of biology. The creation model does not explain the similarities between the basic types (dogs and cats), and that is a serious deficiency, because Darwin already had an elegant explanation for the similarities between taxonomic groups. Creation restricts natural selection and mutation in an arbitrary way. Therefore the dynamic-creation model fails to be a consistent and complete framework for dealing with biological data. It cannot replace common descent. It can be understood only as an attempt to reintroduce the Genesis kinds and not as the result of a genuine attempt to capture the diversity and unity of life. Despite the claim that it is dynamic and modern, and despite the fact that it factually contains more evolution than its formulation reveals, the dynamic-creation model offers no progress beyond Buffon, the eighteenth century French zoologist. It is essentially a pre-Darwinian view of life. And since no real innovative work is done by nature after the divine creation of the basic types (only variation within bounds), it is essentially a static theory. Mark Ridley classified the theory as static, and he was right after all. Behe's irreducible complexity and Dembski's complex specified information likewise are inadequate to explain the similarities we see between, for example, cats and dogs. Dembski has not even made up his mind about the truth of common descent. Without an explanation of the similarities, there is a real gap in their theories of life. So far nobody has produced a full alternative explanation for all the observations that common descent neatly explains. Therefore, it can safely be said that there is currently no alternative to common descent. Common descent is the only nonarbitrary and consistent theory of descent in biology compatible with the evidence.This is a modified and expanded version of chapter 3 in Why Intelligent Design Fails. Notes
Further Reading
|
|
|

|
| Korthof blogspot | home: wasdarwinwrong.com | wasdarwinwrong.com/korthof84.htm |
| Copyright ©G. Korthof 2006 | First published 3 Oct 2006 | Updated: 29 Jan 2014 F.R./Notes 14 NOv 2021 |
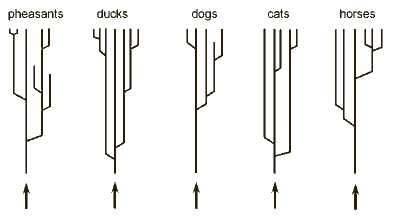
 each genus has many more species than are depicted in the figure. The pheasant family consists of 38 genera and 155 species, the duck family of 41 genera and 147 species, the dog family of 12 genera and 34 species, and the cat family of 37 species.
These numbers dwarf in comparison with families of insects. A beetle family (weevils; see picture) contains around 65,000 species
(Tudge 2000
each genus has many more species than are depicted in the figure. The pheasant family consists of 38 genera and 155 species, the duck family of 41 genera and 147 species, the dog family of 12 genera and 34 species, and the cat family of 37 species.
These numbers dwarf in comparison with families of insects. A beetle family (weevils; see picture) contains around 65,000 species
(Tudge 2000  A general implication of the dynamic-creation model is that all "end products," that is, all species, are not directly created. Since there are no "basic types" alive today, no species we encounter today has been created. The beautiful ornamentation of the Argus pheasant, of which Darwin remarked "was more like a work of art than of nature" (Darwin
A general implication of the dynamic-creation model is that all "end products," that is, all species, are not directly created. Since there are no "basic types" alive today, no species we encounter today has been created. The beautiful ornamentation of the Argus pheasant, of which Darwin remarked "was more like a work of art than of nature" (Darwin