1 |
The origin of DNA, chromosomes and genomes |
| |
Both theories of independent origin claim that all forms of life originated directly from self-assembled genomes. So, let's have a closer look at genomes first.
- All eukaryotic animals have a dual genetic system
Nuclear DNA and mitochondrial DNA (located outside the nucleus). The incorporation of the mitochondrion into an eukaryotic cell is a unique historical accident in the history of life that unites all eukaryota. How could independent origin explain that a dual genetic system originated in all eukaryotes with a non-random distribution of genes over the two genomes as we see it today? Moreover, how could independent origin explain that the distribution of those genes is not random, but similar in all animals and plants? Furthermore, how could independent origin explain that mitochondria are inherited from one parent only (15),(49)?
- All eukaryotic plants have a triple genetic system
nuclear DNA; mitochondrial DNA and chloroplast DNA (both located outside the nucleus). The chloroplast genome is larger than the mitochondrial genome and contains up to 156 genes (61). Thus for each plant, 3 genomes must be assembled and come together in one cell, which is highly unlikely. To complicate matters further: whereas a single fertilization event initiates embryogenesis in animals, in flowering plants two genetically identical gametes from the same pollen grain fertilize the haploid egg cell (chromosome complement n) and the diploid (2n) central cell within the ovule (double-fertilization). The result is a diploid (2n) embryo and a triploid (3n) endosperm (75).
- Eukaryotes with more than 3 genomes
Paramecium is a unicellular Eukaryote with a polyploid macronucleus, one or more diploid micronuclei, mitochondria and some harbour endosymbiotic green algae. Species within the genus Cryptomonas have 4 genomes within the cell: the nucleus, the nucleomorph, the plastid, and the mitochondrial genomes. The plastid genome is circular and contains 118 kilobase pairs. This genome is the direct result of secondary endosymbiosis via a viral phage from a red alga (122). The most extreme example of an organism with multiple genomes is a photosynthetic cryptomonad, which has five genomes! (69).
- The extended genome
Two types can be distinguished: exosymbionts living in the body but outside cells, and endosymbionts living inside cells.
- exosymbionts: an estimated 2 to 4 million genes are embedded in the aggregate genome of an intestinal community of ~500 to 1000 bacterial species (50). That is 100 times as many genes as in our own genome. Recent data show ~150 times larger than the human gene complement. These are microbes living in the human gut. Some 1.5 kilograms of bacteria colonize the human gut (109). Microbes are important for breaking down plant sugars (polysaccharides) that are otherwise indigestible, extracting extra calories from ingested food that could then be used as energy, and synthesize essential amino acids and vitamins. Beneficial gut bacteria can produce anti-inflammatory factors, pain-relieving compounds, antioxidants and vitamins to protect and nurture the body (109).
The genomes of the gut bacteria themselves have been enriched by Horizontal Gene Transfer from marine bacteria (105). Recent work suggests a possible role for the gut microbiota in obesity: gut microbiota promotes obesity by increasing the capacity of the host to extract energy (calories) from ingested food. So they are not superfluous, but can be considered as part of our extended genome. On the other hand, the human genome contains genes that detoxify a dangerous toxin that microbes in our intestines produce (37).
Furthermore, it is not only what bacteria do, but also how the genome of the host reacts to bacteria. For example the host genome contains detoxifying genes which need to expressed in the right tissue at the right time. There are hundreds of genes affected by the establishment of a stable bacterial partnership, including some known to play a role in human responses to bacteria. There is evidence suggesting that the animal-bacteria conversation may be basically the same across evolution. All animals and plants evolved in the background of the presence of huge numbers of environmental bacteria. These genes might be considered the core conserved responses of animals to interactions with bacteria (source).
A second obstacle for the independent origin scenario is that although babies are born sterile (110), microbes are transmitted via mother's milk. Apart from bacteria, in the human gut alone there may be as many as 1,200 distinct viruses (83). See also: Gut bacteria.
A third obstacle may be skin bacteria. On the the insides of human elbows 113 kinds of bacteria (based on 16S ribosomal RNA) were found. The densities are: bacteria under the skin: 1 million bacteria per square centimeter compared with 10,000 from the skin surface. That's a lot of diversity and cell volume. It is not yet clear whether they are beneficial, neutral or harmful. (96).
- Endosymbionts:
The majority of insect species host bacterial endosymbionts that live within their cells, are vertically transmitted from parent to offspring, and have their own genome. Some of these endosymbionts are essential for the survival of the insects (113). See further: symbiosis.
- The diploid genome.
If one has to choose between sexual or asexual reproducing organisms as the first life forms, than one quickly comes to the conclusion that asexually reproducing organisms are immensely more plausible candidates than sexually reproducing organisms. Sexual reproducing individuals need another individual of the opposite sex of the same species to reproduce, while asexual reproducing individuals do not need another individual at all (bacteria,amoebae,sponges,sea anemones,algae). Intimately connected with the existence of two sexes is a long list of properties:
- the diploid state of individuals. Without the diploid state, there would be no Mendelian inheritance (Mendelian dominance and recessiveness; genes are present in pairs; the 3:1 ratio) (67)
- the haploid state of their sex cells (only one allele of a gene present)
- gametes (sex cells) of unequal size
- the fusion of the nuclei at fertilisation (producing the diploid state again)
- the process of meiosis which produces haploid gametes. Meiosis is a highly complicated and non-random process. It must guarantee that a gamete contains not only the right number of chromosomes, but also one and only one copy of each chromosome. One missing or one in duplo will result in serious abnormalities or death.
- that internal and external sex organs of both sexes must be produced during development, but only of one sex in one individual
- that the external sex organs must fit the opposite sex of the same species
- that males and females must be produced in equal proportions
and many other properties. In Schwabe's theory sexual or asexual reproduction makes no difference, in Senapathy's theory sexual organisms arose first!
A very important fact is that the genomes of all higher animals and plants are in a diploid state: 2 set of chromosomes in all body cells. The cause is not difficult to discover: one set is from the mother and the other from the father. So, there must be a father and a mother in the first place. This is one of the main reasons why Independent Birth does not work for diploid organisms (higher animals and plants). It is not a problem for evolution because evolution does not start with diploid organisms. Bacteria are haploid (only one copy of every gene). There has been found only one exception to the rule that animals are diploid: the false spider mite Brevipalpus phoenicis which reproduces by parthenogenesis; the females are haploid (9). Regrettably, it is not a candidate for independent origin because an intracellular parasite is involved!
- The female and male genome
The female genome differs from the male genome in at least four ways (79).
- the Y chromosome endows the male with at least several dozen genes that are absent in the female.
- the incomplete nature of X-inactivation means that at least 15% of X-linked genes are expressed at characteristically higher (but often variable) levels in females than in males.
- a minimum of an additional 10% of genes show heterogeneous X-inactivation and thus differ in expression levels among females, whereas all males express a single copy of such genes.
- the long-recognized random nature of X-inactivation indicates that females, but not males, are mosaics of two cell populations with respect to X-linked gene expression.
If the genomes of sexually reproducing species originate independently, then for each species two genomes, a female and male
genome, must be produced which differ in complicated ways. Not a minor difficulty for the theory of independent origin.
- Chromosomes.
Inside every human cell there is an impressive two metres or so of DNA, which must be organized into a compact form
to fit into the cell nucleus. To do this, cells wrap the DNA around histone proteins (66)
(see Fig. 2). Even bacteria have proteins (called 'condensins') that carry out this condensation. To postulate the spontaneous origin of a complex packaging mechanism without the existence of cellnuclei with certain diameters makes no sense. Apart from that, where do these very specific and highly conserved histone proteins come from? Therefore, DNA alone is not sufficient to create a chromosome.
The spontaneous origin of an eukaryotic chromosome is extremely unlikely.
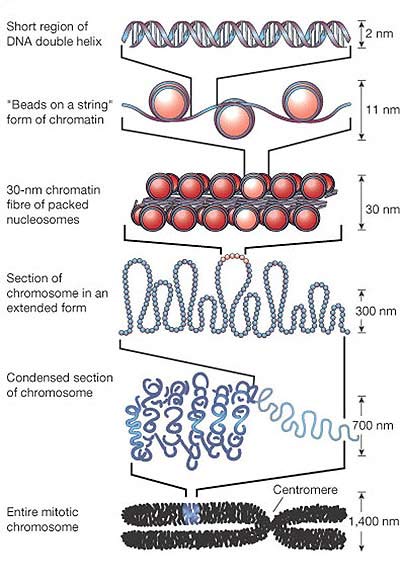
Fig 2. The structure of an eukaryotic chromosome (source).
|
A closer look at chromosomes reveals special characteristics:
- order of genes on a chromosome (synteny = co-linearity of genes). A chromosome segment is conserved when the linear order of genes has been preserved, without interruptions (46, p.138).
"Almost all human genes on chromosome 17 are found on mouse chromosome 11 and "human chromosome 20 appears to be entirely identical to mouse chromosome 2." (47, p.121).
- systematics of karyogram. A karyogram is the appearance of chromosomes in a condensed state (usually cell division) as seen under the microscope. A karyogram enables visual identification of chromosomes. Example: chimp - human karyotype similarity. If genes are randomly aggregated into chromosomes, then there should be no systematics of karyograms. Similar
animals should not have similar karyograms. A karyogram of a crow could be similar to that of a cow.
- sex chromosomes. The sex chromosome pair is a unique pair. Sex chromosomes of all mammals differ in structure and behaviour from the rest of the chromosomes. Females have two X-chromosomes, but one X is inactive (X-inactivation). The X-chromosome is the only inactivated chromosome. It's the exception. How could independent origin explain that systematically only one X-chromosome is inactivated? (62). The sex chromosomes have evolved from a pair of autosomes within the last 300 million years (63). The Y-chromosome is a degenerated X-chromosome. The human Y chromosome evolved from a series of rearrangements of the mammalian X chromosome (87). It lost nearly all DNA of the X-chromosome and has a gene (SRY-gene) that determines the male sexual organs. This was repeated for all mammals just by accident in the primordial pond?
In vertebrates (mammals) all males are XY. In birds the females have two different sex chromosomes (female W-chromosome). It is the systematics of these differences in biological groups, that makes independent origin highly improbably. Sry-gene defines Y-chromosome, but "there are more genes involved in the manufacture of sperm (spermatogenesis) on the mammalian Y chromosome than might be expected by chance". "Are genomes made up of strings of genes in no particular order? It seems not, given the abundance on the mouse sex chromosomes of genes involved in the manufacture of sperm" (8).
- microscopic structure of chromosomes.
Chromosomes need histones for structure (packaging); need proteins for keeping sister chromatids together and for separate them at meiosis. Senapathy and Schwabe considered only naked DNA and only the sequence. This is extreme reductionism. Histones are conserved, they need to be present in huge quantities for a cell to divide. It is impossible to have one cell division without histones. It is very improbably that all these histones are randomly assembled again and again for million of eukaryotic cells, considering the strict constraints for histone sequence. (conserved sequence!). Chromosomes need telomeres (repeats of TTAGGG) at both ends, which refutes the idea that a genome is simply a sequence of DNA. "Cells without TRF2 protein have chromosome abnormalities, disturbed cellcylcus and chromosome ends stick together (48). What is the probability that the telomere repeat sequence ends up at each chromosome end by accident?
- aneuploidy is a condition where whole chromosomes or part of chromosomes are missing. In humans, aneuploidy is the leading cause of birth defects, miscarriage, and mental retardation. Furthermore, aneuploidy is at the roots of cancer (64). This shows a genome must be complete. The problem of distributing chromosomes in the correct way arises at every cell division (meiosis or mitosis). Our bodies are built by many cell divisions.
- origin of chromosomes. Explaining the origin of the DNA sequence (the genome) is not enough. The theory of Independent Birth needs to explain the origin of chromosomes as well as genes and genomes. DNA does not spontaneously organise itself into chromosomes of the higher organisms. Why chromosomes at all? Why do humans have 46 chromosomes in their body cells? Why not 30,000 separate genes floating in the nucleus or one long chromosome containing all the DNA?
|
2 |
Boot problems: how to start an organism? |
| |
How do I start a multicellular organism from a complete but naked genome? (ignoring the generation of the genome itself and ignoring food and energy requirements). Here I present the following fatal development-technical obstacles for the hypothetical independently born organism:
- Overlapping generations

For all mammals, not only does the foetus live within the mother, but also do the foetal germ cells (the potential future generation) live within the foetus (the foetal germ cells appear at 4 weeks). Three generations necessarily overlap in time. The figure is just a snapshot in time. It continues into the past and into the future. There is no point in time that any generation is independent.
Insects: telescoping generations can occur in parthenogenetic species, such as aphids or other life forms that have the ability to reproduce without ovum fertilization. This occurrence is characterized by a viviparous female having a daughter growing inside her that is also parthenogenetically pregnant with a daughter cell.
How, could these organisms originate independently?
- Fertilization
In mammalian zygotes, sperm entry is linked with reorganization of the cytoskeleton, cortical granules, endoplasmic reticulum and mitochondria. Sperm entry also causes changes in zygote shape: formation of the second polar body. In sea urchins, ascidians and frogs, sperm entry triggers cytoplasmic movements, which are associated with successful progress of the establishment of embryo polarity. (117). The absence of centrosome in the unfertilized egg necessitates the delivery of the centrosome from the sperm. Without centrosome the egg does not develop.
- Maternal and paternal load
After fertilization the embryonic genome is inactive in all animals. The fertilized egg contains and uses proteins and RNA from the mother. Only after a number of celldivisions gene expressions of the genome of the embryo starts. For example, the Zebrafish starts transcription of its genome only after 10 division cycli. Human sperm carry some 10 species of RNA, which may provide important signals to the developing embryo (118).
- Necessity of Maternal effect genes in animals and plants for body axes
- Animals:
The founding example of the phenomenon we now call maternal-effect inheritance was discovered in Lymnaea snails. Simply put, it is not the genotype of the individual animal that determines a particular phenotype (in this case the dextral or sinistral shell); rather, it is the genotype of the mother that determines the phenotype of her progeny (104).
The development of multicellular animals depends on the establishment of three body axes. The first two are: head-to-tail (anterior-posterior) and back-to-front (dorsal-ventral) axes. Once these are specified, right and left follow automatically. How are the first two axes constructed? In some animals in the egg, in other animals at fertilization (by the sperm's point of entry), in others still by the unequal distribution of the yolk within the egg (103). Wallace Arthur notes that cell differentiation of the developing embryo leads to an 'infinite regress'. The triggers for differentiation ultimately lead all the way back to the egg (54). In Drosophila the head-to-tail and back-to-front axes, are established in the egg before fertilisation or, in some cases, as an immediate result of fertilisation (16,29). Nobel Prize winner Christiane Nüsslein-Volhard discovered the importance of maternal effect genes to the formation of the body axis in Drosophila (31). The products of maternal effect genes initiate the head-to-tail axis while the egg is still in the egg chamber of the mother (46, p.129). Scott Gilbert (28, p.271) lists 14 maternal effect genes in Drosophila. But also in vertebrates (zebrafish and the frog Xenopus) maternally derived control systems implanted into the egg before fertilisation play a major role in arranging the layout of the embryo's body. For example the dorsal-ventral axis in the zebrafish embryo is controlled by maternal mRNA (82). In humans body axes are not determined by the mother, but the first few cell divisions of the human embryo are under
maternal, not embryonic, control (76, p.49).
- Left-Right asymmetry: the internal body of animals is not 100% symmetrical. This is called Left-Right asymmetry. Humans and other vertebrates have striking asymmetries in the placement of organs such as the heart, lungs, liver and gut. Probably, in mammals the Left-Rright asymmetry is under maternal control also.
- Maturation: the immature egg needs maturation. Maturing mammalian eggs "go through complex changes: they grow and switch many genes on or off in preparation for future development." (27). Also, small mRNA's in the cytoplasma of the Drosophila oocyte have an important function in the suppression of hybrid dysgenesis. This is an example of epigenetic inheritance: the inheritance of information outside the nucleus (98). How can all these maternal effects happen if there is no mother?
- germ cell specification: germ cells can be specified either by maternally inherited determinants (preformation) or by inductive signals (epigenesis). Preformation excludes spontaneous origin because there is no mother. Example organism: Drosophila melanogaster (127).
- Plants:
In plants, it was long thought that any parental effects on embryogenesis were nonexistent. Over the past decade, however, several mutations that exert maternal effects on embryogenesis have been described in the model plant Arabidopsis thaliana. Now also paternal effects on plant embryogenesis have been discovered. A temporal cue provided by the sperm cell triggers the events leading to the first asymmetric division of the plant embryo. These findings suggest that there may be extensive maternal control over early embryogenesis that is mediated, in part, by cytoplasmically stored products (101).
- Necessity of Maternally and Paternally imprinted genes
There is another fatal obstacle for independent origin called imprinting (40). Gene imprinting, the differential expression of alleles of the same gene depending on parent-of-origin, independently evolved in mammals and in flowering plants. Imprinting occurs in the placenta of mammals and the endosperm of plants, structures that nourish the developing embryo.
In animals imprinting of genes in oocytes and spermatozoa is a requirement for normal development. Some maternal genes repress the formation of cells that will form the placenta while some paternal genes stimulate them. Both sets of genes in combination are necessary to guarantee a balance between placenta forming cells and embryo forming cells. Imprinting is based on sex-specific activation of certain genes in the female and others in the male germ cells. This implies that in mammals, without contribution from the male sperm nucleus, it is not possible to initiate normal embryonic development (85, p.98-99).
Imprinting (methylation) is a mechanism to activate or inactivate genes. "The ability to mark a gene as coming either from the mother or the father is called genomic imprinting. Imprinting adds additional information to the inherited genome. That's why it is an epigenetic effect. It also provides a reminder that the organism cannot be explained solely by its genes. In other words: the Sequence is not enough.
It is currently thought that perhaps a few hundred of the 20,000 or so human genes are subject to imprinting, although only 63 are confirmed so far. Particularly striking is the number of imprinted genes that act in the fetal growth pathway mediated by insulin-like growth factor II (IGFII). The other theme that is beginning to emerge is the role for imprinting in brain development (55). Thus, the insulin-like growth factor II Igf-2 gene is methylated in the mouse egg and unmethylated in the mouse sperm. The expression of UBE3A in the brain comes only from the unmethylated maternal derived genes." (28, 5). Failure of imprinting or faulty imprinting causes neurological disorders, abnormal growth and some cancers (78). A defective imprinting of the maternal copy of the UBE3A gene causes the neurodevelopmental disorder Angelman syndrome in humans. Only the maternal copy is active in neurons, the paternal copy of UBE3A is silenced, so humans are effectively haploid for the UBE3A gene. If the maternal copy is inactive Angelman syndrome develops (119). "Mammals have a requirement for a balance between maternally and paternally imprinted genes." (9). "Maternal and paternal genomes are both necessary for normal development in mice, and this is believed to account for the absence of parthenogenesis [development without a father] in mammals." (65, p.11). How does independent origin create maternal and paternal imprinting, when the first cell doesn't have parents at all?
A lesson from cloning animals: developmental reprogramming
So far, there's only one proven method of turning back a body cell's developmental clock: take an adult cell and fuse it with an unfertilized egg cell stripped of its own genetic material (73). So, the egg has the capacity for developmental reprogramming. The factors responsible for this are in the cytoplasm, not in the nucleus. This shows why any theory of the origin of multicellular organisms cannot restrict itself to the origin of a genome. The Sequence is not enough! That is the boot problem. The origin of any genome is hard enough and certainly an animal genome, let alone the required specific boot status.
The reason that many cloned animals die before birth or have health problems later in life, is that in cloning only one diploid genome is used (that is the purpose of cloning) and so there is only one epigenetic imprint and never two different ones. The result is that the expression pattern of hundreds of genes in a cloned animal is wrong (Rudolf Jaenisch). The best method of creating a healthy individual is fusing a haploid genome with a female imprint with a haploid genome having a male imprint. This is not cloning. This is called 'sex'.
|
- Necessity of Maternally derived nucleolus
The nucleolus in the zygote and early mammalian embryo is exclusively maternally inherited and originates from the material that is present in the oocyte germinal vesicle. The necessity of the entire nucleolar complement for successful embryonic development has been shown. Thus, the oocyte nucleolar material is essential for successful early embryonic development in mammals. Nucleoli originating from a somatic cell or even from an embryonic stem (ES) cell cannot substitute for the original oocyte nucleolar material. Furthermore, it is speculated that early protein synthesis uses maternally stored ribosomes and RNAs (94).
- Necessity of the paternal centriole in fertilization
The centriole must be properly transmitted during reproduction in sexually reproducing organisms. The centrosome controls microtubule organization. In order to ensure species continuity, a functional centrosome must thus be inherited each generation. In sexually reproducing organisms, the centriole is most often inherited from the sperm, and a functional centrosome arises in the egg following fertilization. Centrosome numbers must be carefully monitored and controlled – supernumerary centrosomes can have devastating consequences leading to improper chromosome separation and cell death. (123).
- Mechanical requirements for development
The ability of embryonic cells can be beautifully studied in a Petri dish (a kind of primordial pond in the lab). When embryonic stem cells from a mouse are cultured in a Petri dish, they form a random assortment of nerve, muscle and blood cells, but not a mouse embryo. A mouse cannot form in a Petri dish. For a mouse to develop, it is absolutely necessary to have the organizing influences of both the outer trophectoderm cells of the mouse blastocyst and of the mother's uterus after implantation (85, p.100). The same is valid for a fertilized human egg. Despite the fact that the genome is complete, for its realisation, the intense interaction - the symbiosis with a second organism, the mother - is required (85, p.145). Unfortunately, there are no mothers in the primordial pond.
- Structures that make no sense in independently born animals but are encoded in
the genome
In animals extraembryonic tissues develop which protect and feed the growing embryo. The egg of birds contains large amounts of yolk produced by the mother. Without yolk the embryo cannot grow. Blood vessels outside the embryo transport nutrients from the yolk to the embryo. This is a fixed part of development and is under genetic control. It is encoded in the genome. The theory of independent origins is based on the primacy of the genome. In hypothetical independently born birds, there is no yolk, so extraembryonic blood vessels make no sense and do not work.
In mammalian embryos extraembryonic tissues are even more obvious, important and complex. They form a part of the placenta;
the other part is formed by the mother. Another organ is the umbilical cord which guarantees the supply of oxygen and nutrients and provides the passageway for waste products. In this way, the embryo makes use of the mother's organs as long as its own organs cannot function independently (85, p.100).
In the independent scenario, genomes would originate which form pointless structures and which make only sense in animals that are inside an egg produced by the mother (birds) or develop inside a mother (mammals). Yet they are encoded in the genome. Evolution does not have these problems because evolution does not start with birds and mammals.
- Translating the first gene: a vicious circle
 A next boot problem is the vicious circle of translation of the first genes in a fertilised egg (41). In short: DNA needs to be copied to RNA and RNA needs to be translated into protein. Translation requires a dictionary. A dictionary with 64 entries. That dictionary is stored in DNA (it is inherited). However, because it is stored in DNA it needs to be translated first. Now a vicious circle arises: to translate any DNA one needs a dictionary, but the dictionary itself must first be translated to be operational. It looks like an unresolvable boot problem. The only way out is to have a fully translated and operational dictionary ready in the fertilised egg. How? From the individual who produces the egg: the mother. Conclusion: we need a mother as a source of an operational DNA-protein dictionary for the translation of the information of DNA in the fertilised egg.
A next boot problem is the vicious circle of translation of the first genes in a fertilised egg (41). In short: DNA needs to be copied to RNA and RNA needs to be translated into protein. Translation requires a dictionary. A dictionary with 64 entries. That dictionary is stored in DNA (it is inherited). However, because it is stored in DNA it needs to be translated first. Now a vicious circle arises: to translate any DNA one needs a dictionary, but the dictionary itself must first be translated to be operational. It looks like an unresolvable boot problem. The only way out is to have a fully translated and operational dictionary ready in the fertilised egg. How? From the individual who produces the egg: the mother. Conclusion: we need a mother as a source of an operational DNA-protein dictionary for the translation of the information of DNA in the fertilised egg.
- Necessity of learning from parents.

The study of problems with rehabilitation of animals born in captivity (for example in zoos) is extremely useful for understanding the problems of an hypothetical organism originating spontaneously in a hypothetical primordial pond. An individual animal must be able to fly, swim, climb, hunt, search out food, avoid predators, recognize members of its own species and members of the same or other sex in order to survive and reproduce in the wild. This is impossible for birds and mammals (and maybe more animals) without parents. Perhaps the most famous rehabilitated wild animal is the African lioness Elsa, who was immortalized in the movie Born Free. Hand raised and habituated to human "parents", she was taught to be a wild lion again. In the process, she nearly died because she seemed incapable of hunting (108). Despite these rehabilitated animals have a head start in life, they still cannot survive in the wild without learning the necessary survival skills from their parents.
- Necessity of growing up with members of your species
Early social isolation results in adult behavioral and cognitive dysfunction. Juvenile social isolation and neglect influence adult cognitive function and social interactions: brain damage, mental illness.
See Social deprevation.
- Behavioural and sexual imprinting
An example of behavioural imprinting is that ducklings respond to the sight of the first large moving object they see. Under natural conditions this is the mother. This behaviour has survival value. Sexual imprinting for example is that youngsters become imprinted on the image of the parents who care for them, and in adulthood this image is the model for their choice of a mate (80).
 A different boot problem arises later in life. A good example is how songbirds learn their species-specific songs
(song imprinting). Like humans birds must hear the sounds of adults during a sensitive period (10). To sing properly, the young male White crowned sparrow must learn the songs of an adult, or else he will develop highly
abnormal songs (102). All other problems ignored, organisms that arise independently from a primordial pond, don't have a clue of how a parent or mate looks like. If imprinting occurs they would be imprinted by an individual of a different species. Simply because the first individual of a species cannot encounter individuals of the same species by definition. How to find a mate if you don't have a clue what to look for and secondly, if there are no mates at all? The first individual of a species would be doomed to extinction (81). A different boot problem arises later in life. A good example is how songbirds learn their species-specific songs
(song imprinting). Like humans birds must hear the sounds of adults during a sensitive period (10). To sing properly, the young male White crowned sparrow must learn the songs of an adult, or else he will develop highly
abnormal songs (102). All other problems ignored, organisms that arise independently from a primordial pond, don't have a clue of how a parent or mate looks like. If imprinting occurs they would be imprinted by an individual of a different species. Simply because the first individual of a species cannot encounter individuals of the same species by definition. How to find a mate if you don't have a clue what to look for and secondly, if there are no mates at all? The first individual of a species would be doomed to extinction (81).
- Gut bacteria
Mammals:
The mammalian gut contains around 1013 bacteria, the majority of which belong to the phyla Bacteroidetes or Firmicutes. Coevolution with these microbes has driven the functional morphology and immune function of the gastrointestinal tract. Without microbes, aberrant physiology develops together with problems in host defense. Both can be rectified upon reintroduction of bacteria. Additionally, childhood exposure to microbes can direct the maturing immune system to develop a tolerance to environmental antigens ('hygiene hypothesis'). More recently this concept has been extended to include "macrofauna" of the gut, such as helminth parasites. Helminths have been found to be a major force underlying the evolution and selection of interleukin genes (107).
Humans have gut bacteria (see also above "the extended genome"). "At birth, babies emerge from a sterile environment into one that is laden with microbes. The infant's intestine then rapidly becomes home to one of the densest populations of bacteria on Earth.
Numerous benefits arise from our intestinal microbiota: these bacteria help in extracting nutrients from food, and are instrumental in the development of the gut, and the immune system after birth." (91).
Insects: Successful development of fruit flies depends on a gut bacterium that interacts with its host's insulin-signaling pathway. In related species of the wasp genus Nasoni, the gut microbiome and host genome represent a coadapted "hologenome" that breaks down during hybridization, promoting hybrid lethality and assisting speciation (121). A coadapted hologenome is a most severe barrier to independent origin.
Termites. A final example of a boot problem, a serious one, is in termites.
Termites cannot digest wood, however, micro organisms in their gut can. "The gut of wood-feeding termites is a tiny but astonishingly efficient bioreactor, in which microbes catalyse the conversion of lignified plant cell walls to fermentation products that drive the metabolism of their host. Molecular phylogenetic data have revealed the presence of hundreds of microbial species in this microlitre-sized environment" (92). "The insects need to reinfect themselves with their digestive aids in every generation and in some cases after every moult (because the lining of the gut is lost in moulting)" (18).
The termite hindgut contains ~107 prokaryotic cells per µl with over 250 different species of bacteria, and the bacteria themselves contain viruses (bacteriophages) (114). The total of genomes of termite+bacteria+viruses is called metagenome. Here we have an overlap with ecology and adaptation. Before discussing these topics, an important one first: parental care.
|
3 |
The energy costs of parental care: the making of an individual |
| |
 Figure 3.
Figure 3.
Does the very first human individual in the Independent Birth scenario have a navel? If so, where does it come from?
If not, how did the embryo got its oxygen and nutrients?
|
 Figure 4. Figure 4.
A mother suckling her baby symbolises the fact that independent origin of mammals is at least very implausible.
|
|
A mother suckling her baby symbolises the fact that independent origin is impossible for mammals. The umbilical cord and the navel are the symbols for a dependent existence during pregnancy. The female breast is the symbol for a dependent existence during the months after birth.
Internal gestation
All mammals have internal gestation: their embryos develop inside the mother's body. Energy is always required for the growth of the embryo. Birds invest energy in large eggs, mammals invest energy in internal gestation. The whomb is a complex organ. To create an artificial womb "remains in the realms of science fiction" (17). The development of a human fetus shows a range of physiological phenomena: attachment of the embryo to the lining of the womb (endometrium), umbilical cord supplies oxygen, lungs are filled with protective amniotic fluid, at 23 weeks ready to take oxygen, production of a foamy surfactant liquid that stops the lungs from collapsing between breaths, etc. Shortly before birth the baby automatically switches from making fetal hemoglobin to making adult hemoglobin. How can genomes in the primordial pond know that and know when to do that?
Lactation
Nurturing a large-brained baby imposes enormous energy costs on the mother because of the burden of lactation, which is far more costly than gestation. To produce breast milk, females require an extra 500 calories a day. A baby weighing 3.5 kilograms (7.5 pounds) needs about 400 kilocalories a day. An important factor is that the baby needs maintaining its own body temperature after birth and indeed its metabolic rate rises rapidly.
In small mammals, lactation triples the amount of food that must be eaten by a female (68). A newborn elephant calf consumes up to 11 -15 litres of milk in a single day and it may nurse for up to 2 years of age or
older. Mammalian milks have a wide range of dry matter levels, from 8.5% in asses to 64.4% (!) in the northern elephant seal (95). The energy and food requirements of the fetus and baby demand an uninterrupted supply.
Human females store those extra calories in their body in case food is unavailable. Average percentages of fat in young women: 27% and for men: 15% (95).
How does the primordial pond do all that? "Before and soon after birth, temporarily protected by antibodies that our mother has given us through the placenta and in her milk, our immune system prepares for the inevitable onslaught of the many pathogens to come." (53).
Baby teeth (milk teeth) appear from the age of about six months. The newly born baby is helpless and completely depends on its mother for survival. Unlike our close cousins the chimpanzees, humans have a prolonged period of development after weaning, when children depend on their parents to feed them, until at least age 6 or 7. Researchers have known since the 1930s that we take twice as long as chimpanzees to reach adulthood.
Other animals
There is much variation in parental care among animals. Many animal parents do not care for their young, but others provide their offspring with costly, time-consuming services, ranging from protecting them against predators and environmental stresses to giving them shelter or food (106). Therefore, many young animals cannot survive without parental help (external link: overview of Parental Care). Some examples:
- Mammals: lion cubs are born blind and helpless, and are dependent on the mother to survive. A domesticated cat is born blind and helpless; senses develop only after 10-12 days.
"Implantation involves a series of steps leading to an effective reciprocal signalling between the blastocyte (zygote) and the uterus" (32). Implantation is under genetic control. The gene L-selectin is expressed by trophoblast cells of the blastocyst and is a crucial mediator of the initial interaction between the embryo and uterine lining, the first step in implantation (42). How could the presence of this gene be explained in a genome that originated in a primordial pool, without the womb of a mother? If it is not there from the start, there can be no selection for it subsequently.
- Non-placental mammals such as kangaroos have short gestation period, but the young are extremely immature at birth and depend also on mother's milk for further development. Therefore, mammals with short gestation times cannot be born independently in the primordial pond. Even egg-laying mammals, like the duck-billed platypus and the spiny anteater, nourish their young with milk.
 Nearly all birds incubate their eggs. An egg is relatively independent of the mother bird when compared to mammals, because it contains a food store (yolk). An extreme case of high energy investment in an egg is New Zealand's kiwi, whose eggs (at 400 - 435 g, roughly 20% of the female's weight) provide nutrition for one of the longest incubation periods (70-74 days), resulting in a fully feathered chick with adult-type plumage (95). All eggs need incubation. A constant temperature (higher than the temperature of the environment!) is crucial for survival. There is one exception: the 22 species of megapodes are sometimes known as thermometer birds or incubator birds because of their habit of laying eggs in the ground and then leaving them. The eggs are incubated by volcanic heat, the heat of sand or the warmth from decaying vegetation. However, these spots need to be carefully selected. Nearly all birds incubate their eggs. An egg is relatively independent of the mother bird when compared to mammals, because it contains a food store (yolk). An extreme case of high energy investment in an egg is New Zealand's kiwi, whose eggs (at 400 - 435 g, roughly 20% of the female's weight) provide nutrition for one of the longest incubation periods (70-74 days), resulting in a fully feathered chick with adult-type plumage (95). All eggs need incubation. A constant temperature (higher than the temperature of the environment!) is crucial for survival. There is one exception: the 22 species of megapodes are sometimes known as thermometer birds or incubator birds because of their habit of laying eggs in the ground and then leaving them. The eggs are incubated by volcanic heat, the heat of sand or the warmth from decaying vegetation. However, these spots need to be carefully selected.
Parental care: over 90% of living birds have biparental care. A small group of species have exclusively female care, but less than 100 species of living birds have exclusively male parental care.
- Parental care in reptiles, amphibians, fish, arthropods.
 Amphibians: 18% salamanders and 6% frogs and toads exhibit some sort of parental care. At least 25 % of fish demonstrate parental care.
Even dinos had parental care. There's plenty of evidence to suggest that dinos baby-sat their offspring. A predatory dinosaur called Oviraptor, for example, was discovered in the Gobi Desert in 1993, its fossilized remains protecting a brood of eggs (Science). Even scorpions, cockroaches, wasps, bees, ants, beetles, salamanders, lizards, frogs, termites and sea cucumbers have parental care (11).
Fishes built nests and care for their eggs (mouthbrooders: cichlids). Red king crab adult females brood thousands of embryos underneath their tail flap for about a year's time. When the embryos are fully developed they hatch as swimming larvae (12). Females crabs hold eggs in a "brood pouch" between their wide abdomen and their cephalothorax (13). Sticklebacks, pipefish, and seahorses belong to the same order of fish, Gasterosteiformes, and are noteworthy in the animal kingdom for evolving highly advanced system of male care. Male Pipefish incubate young in a brood patch. Male seahorses have evolved elaborate vascularization in the pouch where nutrients are transferred to the eggs much like a female mammal transfers micronutrients across the placenta (14). Another example of parental care is the octopus:
"Once the eggs have been laid, the female octopus will gently caress the eggs with her suckers to keep algae and bacteria from growing on them. She keeps the eggs oxygenated by gently squirting them with streams of water from her syphon. After the developing octopus turns in the egg so the tip of it's mantle is at the unattached end of the egg and is ready to hatch. The female's gentle caresses become more violent to help the baby octopus escape from the egg case. Most females will not eat after laying eggs and die soon after her eggs have hatched." (19).
Amphibians: 18% salamanders and 6% frogs and toads exhibit some sort of parental care. At least 25 % of fish demonstrate parental care.
Even dinos had parental care. There's plenty of evidence to suggest that dinos baby-sat their offspring. A predatory dinosaur called Oviraptor, for example, was discovered in the Gobi Desert in 1993, its fossilized remains protecting a brood of eggs (Science). Even scorpions, cockroaches, wasps, bees, ants, beetles, salamanders, lizards, frogs, termites and sea cucumbers have parental care (11).
Fishes built nests and care for their eggs (mouthbrooders: cichlids). Red king crab adult females brood thousands of embryos underneath their tail flap for about a year's time. When the embryos are fully developed they hatch as swimming larvae (12). Females crabs hold eggs in a "brood pouch" between their wide abdomen and their cephalothorax (13). Sticklebacks, pipefish, and seahorses belong to the same order of fish, Gasterosteiformes, and are noteworthy in the animal kingdom for evolving highly advanced system of male care. Male Pipefish incubate young in a brood patch. Male seahorses have evolved elaborate vascularization in the pouch where nutrients are transferred to the eggs much like a female mammal transfers micronutrients across the placenta (14). Another example of parental care is the octopus:
"Once the eggs have been laid, the female octopus will gently caress the eggs with her suckers to keep algae and bacteria from growing on them. She keeps the eggs oxygenated by gently squirting them with streams of water from her syphon. After the developing octopus turns in the egg so the tip of it's mantle is at the unattached end of the egg and is ready to hatch. The female's gentle caresses become more violent to help the baby octopus escape from the egg case. Most females will not eat after laying eggs and die soon after her eggs have hatched." (19).
Even simple marine organisms as starfish and sea cucumber show parental care.
"Some brittle stars release gametes into bursal slits in which they brood their embryos; cilia circulate ocean water in these surface slits, facilitating excreation and respiration. Numerous sea cucumbers brood young among tentacles, in special pockets in their body walls, or in the coelom or gonad, where fetilized eggs have been retained." (59, p.321)
 Viviparious insects: the Hippoboscoidea fly superfamily has a unique and unexpected property: viviparity (to give birth to live young rather than laying eggs), internal developing larvae, and adaptation of the female accessory gland to function as a nutrient synthesis and delivery system. Very much like pregnancy and lactation in mammals! Viviparious insects: the Hippoboscoidea fly superfamily has a unique and unexpected property: viviparity (to give birth to live young rather than laying eggs), internal developing larvae, and adaptation of the female accessory gland to function as a nutrient synthesis and delivery system. Very much like pregnancy and lactation in mammals!
- Plants: mammals have a placenta, plants have seeds. "Most of a seed's food reserves are transferred directly from maternal to offspring tissues after fertilization. These food reserves have a major influence on seedling fitness. Seeds with larger food reserves produce more vigorous seedlings. Larger seeds are a greater metabolic cost to the mother." (65). Independently born plant genomes would not have the benefit of a private food source (a rather lethal condition),
but the genome itself would contain all the genes to produce seed for the next generation? by accident?
- Anisogamy: female gamete is bigger than a male gamete because it stores nutrients for the embryo (food-storing gamete). Explanation for Anisogamy is that eggs are costly to produce and sperm is cheap (49). If it is costly, it means it is highly unlikely that eggs arose out of building blocks. Only plants can live from solar energy.
Conclusion: the existence of parental care, internal gestation, lactation (in mammals), child care (many of the higher animals) is a strong refutation of Independent Origin, because a constant supply of food is necessary in all stages from egg formation through embryo development to child development. This alone is sufficient to refute independent origin. We all live, because our parents invested time and energy in us. There is no substitute for the womb and parental investment.
Parental care and altruism
Altruism is helping others at a cost for oneself. Altruism is directed preferentially to genetically related individuals. To regulate behaviour conditionally in response to different degrees of kinship, organisms require mechanisms to discriminate genetic relatedness. Such mechanisms have been discovered in a variety of nonhuman species ranging from social amoebas, social insects, and shrimp, to birds, aphids, plants, rodents and primates. Humans have an evolved system for detecting genetic relatedness, a human kin detection system, coupled to two output systems: one regulating altruism, the other regulating mate choice (89). The theory of independent origin must fail to explain the origin of altruism because the first individual of a species neither has a mother nor siblings. On the theory of special creation of the first human pair, incest among the children cannot be avoided.
|
| 4 |
Adaptation and Ecology: how to start an ecosystem? |
| |
One cannot start an ecosystem with top-predators (lions), because they depend on herbivores (zebras).
One cannot start with herbivores either, because they depend on plants (grasses).
One cannot start with parasites, because they need hosts.
One cannot start with bees, because they depend on flowers.
Without plants planet earth is uninhabitable. Plants transform solar energy, water and atmospheric CO2 into plant carbohydrate matter by photosynthesis. This plant matter then sustains the global food web and becomes the source of food for animals. In this paragraph I mention a few examples of adaptation and ecology that make independent origin and survival technically impossible.
- Parasites: more than half the world's species live in or on the bodies of other organisms.
Some 80 percent of insects are parasitic on animals and plants (86). This makes independent origin for those species much and much more difficult, if not impossible. Organisms are not isolated individuals that could live without other organisms. In his ecology textbook "Essentials of Ecology", Colin Townsend (44) devotes a fascinating chapter to 'Organisms as Habitats' The whole chapter reads as a refutation of independent origin. Obligatory reading for Senapathy and Schwabe. If the creators of the theory had read this chapter before they published their theory, they would have abandoned their theory immediately.

louse |
Parasites completely depend on their host for survival (34). One species of louse lives exclusively on the human head (see illustration). It needs blood from the host 3-4 times a day to survive.
"The malaria parasite Plasmodium falciparum leads a complicated life, existing mainly inside liver cells and red blood cells in human host and, when residing in mosquitos (notably Anaopheles gambiae), being associated with the insect's gut and salivary glands" (24). Female mosquitos need blood from reptiles, birds or mammals for the growth of their eggs. There are 3,000 mosquito species (Culicidae). They all need blood for their eggs. For the theory of independent origin in the strict sense it means that 3000 different individuals arising from the primordial pond need to find a suitable animal to reproduce.

Fig. 5. Anopheles. |
The lives of humans and mosquitoes have become intricately intertwined. The mosquito's ability to exploit almost any type of water - natural ponds and marshes or human creations such as irrigation ditches and used tires - is testimony to its evolutionary ingenuity. Spielman believes that the suite of enzymes produced by particular species of mosquitoes is closely tailored to the biochemistry of its chosen hosts. Anopheles gambiae thus probably produces enzymes that work best against the clotting and inflammatory biochemical pathways of its preferred target: humans (22).
 To say that the malaria-carrying mosquito Anopheles gambiae is well adapted to its role as a human parasite is like saying that Pavarotti is a pretty good singer.
The common house mosquito, Culex pipiens, implicated recently as a major vector of West Nile virus in the United States, feeds at night and strikes birds, livestock, and people; it appears to be attracted generally by heat and carbon dioxide. There is also a high degree of feeding specialisation. Among the seven are some (such as Anopheles gambiae "proper") that concentrate almost exclusively on humans and others (such as Anopheles quadriannulatus) that feed almost exclusively on animals. Those specialisations are controlled by genes and therefore are parts of their genomes. To say that parasite and host are well adapted implies that their genomes are well adapated. Their genomes contain genes that would not be present and can only be explained by the presence of the other organism. To say that the malaria-carrying mosquito Anopheles gambiae is well adapted to its role as a human parasite is like saying that Pavarotti is a pretty good singer.
The common house mosquito, Culex pipiens, implicated recently as a major vector of West Nile virus in the United States, feeds at night and strikes birds, livestock, and people; it appears to be attracted generally by heat and carbon dioxide. There is also a high degree of feeding specialisation. Among the seven are some (such as Anopheles gambiae "proper") that concentrate almost exclusively on humans and others (such as Anopheles quadriannulatus) that feed almost exclusively on animals. Those specialisations are controlled by genes and therefore are parts of their genomes. To say that parasite and host are well adapted implies that their genomes are well adapated. Their genomes contain genes that would not be present and can only be explained by the presence of the other organism.
 | .jpg "New World screwworm fly")
New World screwworm
©wikipedia |
The New World screwworm fly Cochliomyia hominivorax is a species of parasitic fly that is well known for the way in which its larvae (maggots) eat the living tissue of warm-blooded animals. New World screwworm flies are dependent on a host (obligate parasitism), as their larvae require living tissue for food.
- minimal genomes. Viruses are a splendid example of parasites with a minimal genome. They cannot 'live' on their own and most importantly, need hosts to reproduce. Their genomes are reduced to an absolute minimum. Parasitic bacteria have adapted to life in animal hosts to such an extent that they have lost numerous genes as they became dependent on materials supplied by their hosts (7, 60). Some of these parasites, like Mycoplasma and Borrelia, depend on the host to provide key metabolites. Others, like Rickettsia and Chlamydia, are energy parasites that literally steal ATP from the host cell (7).
- symbiosis.
In 80% of land plants, roots associate with the soil fungi Glomeromycetes, forming chimeric organs called Mycorrhizae. Plants depend on Glomeromycetes for mineral resources, especially phosphorus, which is limiting in many soils. On the other hand, the fungi get glucose (made by photosynthesis) from plants (115).
Lichens are splendid examples of symbiotic associations between fungi and photosynthetic microbes such as algae (59). Mycorrhizae are fungi that form a mutual beneficial association with plants: minerals and carbohydrates (made by plants) are exchanged (38), (20). "Rhizobia have specific and intimate interactions with eukaryotic cells: they adhere to plant roots, invade root cells" (36). There is a wellknown symbiosis between nitrogen-fixing bacteria and legumes (Nitrogen availability is limiting to plant growth).
Gut bacteria (E.coli) came by accident in our guts? Special bacteria in the gut of honeyguide bird allow them to digest the beeswax.
Attine ants, which include leaf-cutter ants that can defoliate a tree in one night, can't digest plant matter themselves. But they retrieve leaves and other detritus from their surroundings and heap them up in their nests as offerings for hungry fungi. Thus nourished, the fungi send out nutrient-filled threads that are eaten by their faithful keepers (43).
Examples from insects show that symbioses can result in specialized organs with unique development, innovations in metabolic capabilities that allow new lifestyles, defenses against natural enemies and other environmental challenges, constraints on evolutionary range, and ongoing acquisition of novel genes and capabilities (90).

The mealy bug harbors an endosymbiotic
bacterium that harbors an endosymbiotic
bacterium (116). |

Blue-ringed octopus harbors bacteria in the salivary glands that produce a deadly toxin. The bacteria are transmitted to the next generation by the egg. |
Nested endosymbiosis:
Independent origin and chance symbiosis is even more difficult when three organisms are involved in symbiosis: a heat-tolerant grass in 65°C Yellowstone soils can grow only when infected by a fungus, which is protective only when it itself is infected by a mycovirus (88).
Two bacteria that generate amino acids for their insect host, the mealybug, live one inside the other in an nested arrangement.
One bacterium lives in the cytoplasma of the other. The host has only 121 protein-coding genes. Each of the three participants does not have the complete set of genes to make all necessary amino acids, but together they do (116).
Conclusion: When species A and B depend on each other ecologically, and A and B can arise at any time during the existence of the primordial pond, the probability that they arise at the right time and at the same place, is much lower than when one of the two has an autonomous way of life.
- food:
 Lions prey on zebras, and zebras eat grass. This common sense knowledge would be enough to withhold most people from postulating independent origin of lions, zebras and grass or more generally of carnivores, herbivores and plants. When the very first lion ate the very first zebra, a species got extinct, because there was only one zebra. All organisms need food! Even if independent birth of a complex organism were possible, an animal would die within a few days. Large animals (elephants, whales, dinosaurs) need huge amounts of food. For animals, food means another organism. Does DNA 'know' to start with producing plant genomes, next the genomes of herbivores and finally the genomes of carnivores? Of course there cannot be such knowledge in DNA molecules. A lot more can be said. One example. Insects depend on plants. Each of the world's 300,000 plant species is a target for attack from a range of nearly 400,00 species of plant eating insects (33). How does DNA know that plants have to be produced before insects?
Lions prey on zebras, and zebras eat grass. This common sense knowledge would be enough to withhold most people from postulating independent origin of lions, zebras and grass or more generally of carnivores, herbivores and plants. When the very first lion ate the very first zebra, a species got extinct, because there was only one zebra. All organisms need food! Even if independent birth of a complex organism were possible, an animal would die within a few days. Large animals (elephants, whales, dinosaurs) need huge amounts of food. For animals, food means another organism. Does DNA 'know' to start with producing plant genomes, next the genomes of herbivores and finally the genomes of carnivores? Of course there cannot be such knowledge in DNA molecules. A lot more can be said. One example. Insects depend on plants. Each of the world's 300,000 plant species is a target for attack from a range of nearly 400,00 species of plant eating insects (33). How does DNA know that plants have to be produced before insects?
Big animals need huge daily amounts of food, but even small animals (hummingbird) cannot survive a day without food. Giant pandas are famous for their love of bamboo, but it seems that they also have a preference for old forests (111). Almost all animals need an ecosystem made up of many species to survive. Essentially, we have here a boot problem: how to boot an ecosystem?
- hermit crabs
 have a soft, curled belly that squeezes right into the twists and turns of a snail shell. Inside the shell, the hermit is protected.
Which was first produced by the primordial pond: the hermit crab or the gastropod? Both at the same time? Then the hermit crabs must wait until the gastropod died and the shell becomes available. Since DNA cannot know about what it produces, all interactions between organisms must be cheer luck.
have a soft, curled belly that squeezes right into the twists and turns of a snail shell. Inside the shell, the hermit is protected.
Which was first produced by the primordial pond: the hermit crab or the gastropod? Both at the same time? Then the hermit crabs must wait until the gastropod died and the shell becomes available. Since DNA cannot know about what it produces, all interactions between organisms must be cheer luck.
- internal adaptation. Another example is molecular signalling between embryo and uterus to enable implantation (all mammals). This cannot be 'tested' on day 1 of their existence. If it fails, it means extinction.
- sex organs. Male and female sex organs are an example of reciprocal adaptation. Sex organs are not necessary to survive, but are necessary for reproduction. Sex organs are based on genes, therefore both set of genes must be present in the genomes of at least two individuals of the same species. A one-step selection process would clearly fail, because it means extinction. Either more steps are needed, or more individuals of the 'same species' are needed. More steps means more generations and without well-matched sex organs, there is no next generation. More individuals with exactly the same genome is highly improbable.
- rudimentary eyes. Cave dwelling animals are species that live under ground (mole): blind but with rudimentary eyes. The genes responsible for the behaviour, anatomy and rudimentary eyes of the mole must be present in the same genome. Rudimentary eyes is an example of a set of adaptions of cave dwellers. Some cave dwellers live permanently underground and cannot survive outside the cave environment. These facts are difficult to understand from an independent origin perspective.
- extremophiles.
For every extreme physical condition investigated (high, low temperature, drought, salinity, pH, pressure), extremophilic
organisms have shown up that not only tolerate these conditions but often require them for survival. Their current habitats are not compatible with one primordial pond, because these conditions are mutually exclusive. The same pond cannot be frozen and boil or have high and low acidity at the same time. Strict (obligate) extremophiles cannot live outside their extreme environments. For example a thermophilic bacterium grows at 121°C very well, but cannot grow at all below 85°C (84 p.179). These extreme adaptations are reflected in their genomes. Those genes must be present right from the start.
- oxygen. The simultaneous origin of strict anaerobes (which are killed by oxygen) and
aerobic organisms (which require oxygen) from the primordial pond is technically impossible. Either the obligate anaerobes are killed at current oxygen levels (21%) or aerobic species die at low (or zero) oxygen levels. The level of atmospheric oxygen was too low to support aerobic organisms before about 2.3 billion years ago (72). Any theory of the origin of life must take this into account. If the origin of life is situated at 3.5 billion years ago (as Schwabe does), those forms of life could not have depended on oxygen. They must have been anaerobe. Therefore, no animal cell could have originated and survived at that time.
There is an additional lesson from oxygen: its source. The sources of oxygen are organisms themselves, not geological events. The source of oxygen is photosynthetic organisms. This necessitates a particular chronological order of the origin of organisms: first oxygen-producing organisms, followed by oxygen-consuming organisms. This adds up to the double reason why animals and plants could not originate at the same time: (1) animals could not survive, (2) plants needed to construct a niche for animals: niche construction. Niche construction is a powerful and general ecological argument against independent origin.
- mimicry.

photo: Peter Clausen |
Plants mimic insects and insects mimic plants. Picture shows a praying mantis which mimics an orchid. Other examples are the dead leaf mantis mimicking a dead leaf, the fly orchid, leaf-litter frogs (genus Megophrys) and many insects mimicking leaves. See more pictures here. Independent birth can only explain mimicry as an accidental outcome of random genome assembly. Not very likely and not very helpful.
- seed germination and dispersal. The seeds of some plants actually require passage through an
animal gut in order to germinate. For instance, one African melon species is so well adapted to being eaten by a hyena-like
animal called the aardvark that most melons of that species grow on the latrine sites of aardvarks. Countless other plants
have fruits adapted to being eaten and dispersed by particular species of animals (52).
- plant defences. Plants with thorns: thistle with prickly leaves, rose with thorns on the stem. It makes only sense when predators are present. "To protect themselves, plants have in their armoury of passive defence mechanisms such as strengthened cell walls and antimicrobial compounds, as well as active healing responses." (39). Plants have an immune system. How do plant genomes know that they have to protect themselves against microbes, when they do not have any experience?
- normal development. Gut-living bacteria influence normal development in vertebrates (21).

Fig. 6. Phalaenopsis (orchid) |
epiphytes are plants that grow on the branches of trees and are a common feature of tropical forest canopies. Well-known examples are orchids (Fig 6) and mistletoes. They depend on their hosts to survive. It would be extremely improbable that epiphyte and host (together with a complete rainforest?) arose at the same time and place in the primordial pond. And how do they move to the correct region on the earth?
- venomous snakes and frogs: why genes for poison in independently born genomes at all? accidentally?
toxic for what species? (4). Species are immune to their own toxins. Toxin producing and toxin immunity genes necessarily must be present in the same genome.
- brood parasitism.
 Cowbirds and cuckoos are nest parasites that infiltrate a nest of another bird species, remove one of the eggs, and replace it with their own egg. Nest parasitism enables cuckoos to have their young raised by host birds and frees them from the costs of parenting. The most remarkable thing about cuckoo parasitism is the birds' ability to match the eggs of the host species in size, spottedness, background color, and darkness. Imagine the first independently born cuckoo: she has to find a nest of only five species (commonly parasitized in Britain). Is there a suitable nest available? What if he/she is the first bird born in the primordial pond and no nests are available? End of story.
Cowbirds and cuckoos are nest parasites that infiltrate a nest of another bird species, remove one of the eggs, and replace it with their own egg. Nest parasitism enables cuckoos to have their young raised by host birds and frees them from the costs of parenting. The most remarkable thing about cuckoo parasitism is the birds' ability to match the eggs of the host species in size, spottedness, background color, and darkness. Imagine the first independently born cuckoo: she has to find a nest of only five species (commonly parasitized in Britain). Is there a suitable nest available? What if he/she is the first bird born in the primordial pond and no nests are available? End of story.
There are 136 cuckoo species. "The Common Cuckoo is the best known brood parasite in the Old World, and its wonderful trickery and the counter-defences of its hosts provide clear evidence of an evolutionary arms race." (45). A co-ordinated set of characteristics exists: migratory behaviour, egg size, body weight, diet, habitat, (6), preferences, incubation period, killing of chicks of host, begging call (93). Three conclusions: 1) The very existence of brood parasitism means that egg incubation is costly.
Does the primordial pond offer it for free? 2) If the cuckoo came out of the primordial pond before other (suitable) bird species, it was doomed to extinction. 3) The co-evolutionary arms-race modifies both host and brood parasite. All this is a very strong case against independent origin.
- intracellular parasites. Intracellular parasites need cells to survive. Host and intracellular
parasite originated at the same time in one step? (7).
- Co-adaptation.
 The vast majority of flowering plants - 290,000 species, including coffee, watermelons, cucumbers and sunflowers- depend on animals to spread their pollen (56, p.194). Forests, prairies, meadows, seashores and wetlands all depend on a diverse and healthy community of pollinators. Examples of pollinating animals are ants, bees, wasps, beetles, flies, butterflies, moths, birds and bats. Honeybees (Apis mellifera) are used for the fruit and seed production of more than 100 crops in the United States. There is an intimate connection between plants, their pollinators, our food.
Not a state of affairs that could arise by independent origin.
The vast majority of flowering plants - 290,000 species, including coffee, watermelons, cucumbers and sunflowers- depend on animals to spread their pollen (56, p.194). Forests, prairies, meadows, seashores and wetlands all depend on a diverse and healthy community of pollinators. Examples of pollinating animals are ants, bees, wasps, beetles, flies, butterflies, moths, birds and bats. Honeybees (Apis mellifera) are used for the fruit and seed production of more than 100 crops in the United States. There is an intimate connection between plants, their pollinators, our food.
Not a state of affairs that could arise by independent origin.
Insect pollination brings obvious benefits to each participant. On the other hand there are phytophagous insects using plants
for food. The plants protect themselves by an array of toxic chemicals. Some insects uniquely tolerate the toxic substances of one plant species (99). The holstein butterfly has become one of the text book examples of co-evolution (100).
- In Australia, crabs are the chief planters and propagators of mangrove trees (55, p.285).
- Red-cockaded woodpecker (Picoides borealis) requires pine forests at least eighty years old. Each group requires an average of 86 hectares of woodland to produce an adequate harvest of insect prey. To nest, they hollow out cavities in living, mature longleaf pines eighty to one hundred and twenty years old, in which the heartwood has already been destroyed by fungus (58). The woodpecker in figure 1 symbolises this woodpecker. When species A ecologically depends on species B, and they can arise at any time during the existence of the primordial pond, the probability that they arise in the right order is reduced compared with the case that A and B have no ecological relation.
- timing of behaviour: (timing is crucial!)
- daily rhythm.
Circadian rhythms (from 'circa'=about, 'dies'=a day) persist with a period close to 24 hours, even in the absence of environmental time cues. The circadian clock regulates many aspects of metabolism, physiology and behaviour, in cyanobacteria, plants, fungi and animals including humans. This rhythmicity has been observed right down to the cellular and molecular level, with nearly every cell in the human body showing approximately 24-hour (circadian) oscillations in gene expression (125).
In animals the circadian clock is located in a tiny part of the brain called the suprachiasmatic nucleus and is responsible for a normal sleep-wake cycle (humans: close to seven hours sleep). The rhythm is based on a small number of "clock genes" or "circadian genes" (for example prokineticin-2). Furthermore, the 24-hours periodicity is temperature compensated. If genomes arose directly from the primordial pond, is it just a lucky accident that the genetically encoded cycles match the rotation cycle of our planet? If genomes arose from the primordial pond 620 million years ago, their 24 hours clock would mismatch the length of the day, since the length of one day was about 21.9 hours at that time. The subject is also known as 'chronobiology' or 'biological clocks' (70).
- animal migration by air (birds, butterflies, locusts), by land (wildebeest, zebras, gazelles,
springbok, bison, elk, pronghorn), and by sea (whales, sea turtles, salmon).
 Migration is an adaptive trait, hardwired into a species' DNA and vital to its survival. Famous example: Monarch Butterfly (wiki). Birds are able to use the magnetic field of the earth (wiki). Birds migrating at night use the behaviour of stars to navigate (indigo bunting). How do the first organisms know the geography of the earth or the stars? How do they navigate? How do they know where and when to go? All this knowledge is in their genomes by accident? Extremely unlikely! The theory of independent birth does not predict that organisms should migrate anyway. It predicts organisms adapted to local conditions. Migratory routes and wintering grounds are traditional and learned by young during their first migration with their parents. What if you don't have parents? (first organisms). Some species show circuitous migratory routes that reflect historical range expansions and are far from optimal in ecological terms. An example is the migration of continental populations of Swainson's Thrush, which fly far east across North America before turning south via Florida to reach northern South America; this route is believed to be the consequence of a range expansion that occurred about 10,000 years ago (wiki). If a species originated by independent birth, it has no history by definition.
Migration is an adaptive trait, hardwired into a species' DNA and vital to its survival. Famous example: Monarch Butterfly (wiki). Birds are able to use the magnetic field of the earth (wiki). Birds migrating at night use the behaviour of stars to navigate (indigo bunting). How do the first organisms know the geography of the earth or the stars? How do they navigate? How do they know where and when to go? All this knowledge is in their genomes by accident? Extremely unlikely! The theory of independent birth does not predict that organisms should migrate anyway. It predicts organisms adapted to local conditions. Migratory routes and wintering grounds are traditional and learned by young during their first migration with their parents. What if you don't have parents? (first organisms). Some species show circuitous migratory routes that reflect historical range expansions and are far from optimal in ecological terms. An example is the migration of continental populations of Swainson's Thrush, which fly far east across North America before turning south via Florida to reach northern South America; this route is believed to be the consequence of a range expansion that occurred about 10,000 years ago (wiki). If a species originated by independent birth, it has no history by definition.
- timing of egg-laying:
 Egg laying is occurring earlier in the year in a wide variety of temperate-zone birds. Changes in laying date are consequential because birds (great tits) must time reproduction to coincide with seasonal pulses in resources (caterpillar). If prey advance their emergence more rapidly than birds advance their laying, decreased reproductive success and population decline may result. Timing of egg laying in temperate-zone birds has several cues: day length (photoperiod) begins the process, and then supplemental cues such as food, temperature, or the phenology of other organisms can be used to fine-tune the timing of laying to year-to-year variation in spring conditions.
Egg laying is occurring earlier in the year in a wide variety of temperate-zone birds. Changes in laying date are consequential because birds (great tits) must time reproduction to coincide with seasonal pulses in resources (caterpillar). If prey advance their emergence more rapidly than birds advance their laying, decreased reproductive success and population decline may result. Timing of egg laying in temperate-zone birds has several cues: day length (photoperiod) begins the process, and then supplemental cues such as food, temperature, or the phenology of other organisms can be used to fine-tune the timing of laying to year-to-year variation in spring conditions.
- timing of molting (moulting):
It takes a lot of energy to build new feathers. Molting is, therefore, often timed to coincide with periods of less
strenuous demands, such as after nesting or before migration. (source)
|
| 5 |
Summary and Conclusion |
| |
| method of origin | individual starts with | DNA |
|---|
| independent origin | individual starts with a cell, has no parents, has no ancestors | random origin of every genome |
| creation | individual starts as an adult, has no parents, has no ancestors | all DNA is functional because designed |
evolution
(not: origin of life) | individual starts as fertilized egg, has parents, has ancestors | functional and non-functional DNA |
Let us eliminate the impossible, and see what is left. Let us start with the most complex forms of life and work our way down to the simplest forms of life.
- > It is hard to see how animals with internal gestation and parental care could possibly originate independently in a primordial pond and survive. Internal gestation is the supreme example of dependence on previous generations. Anyway, animals are not good candidates for independent origin and survival, because they ultimately depend on plants, they are heterotroph (74).
This is the argument from ecology. The argument from ecology is a relative argument: survival depends on the timing of the origin of other species. This is a huge problem: more than half the world's species live inside or on the bodies of other organisms. Plants utilising solar energy (they are autotroph 74 ) could be candidates because they at least do not depend on other species for food. Autotrophs are organisms that can grow using carbon dioxide (CO2) as their sole source of carbon. Autotrophs generate the biomass on which all other organisms -including humans- thrive.
- > From a developmental point of view, multicellular individuals (all animals and plants) have boot problems which prevent independent origin. This is an absolute argument: development depends on internal factors.
- > From a genetic point of view, multicellular animals and plants are diploid.
It is inherently implausible that a diploid organism would originate in a primordial pond. That eliminates all diploid animals and plants. (Therefore, the primordial pond illustration (Fig 1) is far too optimistic, because there would be no adult animals and plants in the first place).
- > From a cytogenetic point of view, all animals and plants possess chromosomes, which are structures that go beyond pure DNA. This is another reason to eliminate plants and animals. The genomic point of view, produces additional reasons to eliminate plants and animals from the list of candidates, because animals have two genomes and plants have three genomes.
- > That leaves fungi (eukaryotes) and prokaryotes (mainly bacteria) as potential candidates for independent origin. Eukaryotes are the result of a merger of two life forms ('symbiosis') involving a prokaryote, therefore almost by definition eukaryotes are not candidates for independent origin. The merger is now generally accepted by biologists as a historical fact. Consequently, not only plants and animals, but also fungi can be eliminated as candidates for independent origin. An additional reason to eliminate fungi is that they digest living or dead tissue (heterotrophy).
- > What is now left are (single-celled) haploid prokaryotes (bacteria) as candidates for independent origin. Since Senapathy rejects the independent origin of prokaryotes, there are no candidate species left in his theory. In Schwabe's theory, some prokaryotes are left. A subset of prokaryotes are parasites or symbionts (for example gut bacteria). Parasites and symbionts by definition depend on other organisms (usually higher organisms), consequently fail as candidates for independent origin.
- > What is left are non-eukaryotic autotrophs: cyanobacteria, purple and green bacteria, and also some bacteria and Archaea that do not obtain energy from light (they don't need biological compounds produced by other species, and use inorganic molecules). The photosynthetic prokaryotes can be eliminated as candidates for independent origin, because photosynthesis requires complex and highly evolved proteins (30), (126). However, there are simpler molecules that harvest light energy: retinal pigments of Haloarchaea are simpler than the more complex chlorophyll-based photosynthetic machinery. The haloarchaeal purple membrane constitutes one of the simplest known bioenergetic systems for harvesting light energy (130).
- > So, we are left with autotrophic non-photosynthetic prokaryotes as the only organisms that could originate spontaneously from chemical building blocks and survive. The first organisms must have been autotrophic, because they could not have relied on other organisms for their energy (71), unless enough abiotically produced energy-rich molecules and building blocks were available. This is an open question.
- > The first organisms could neither have generated energy from oxygen, because 3.5 billion years ago there
was no oxygen in the atmosphere. Oxygen is a by-product of photosynthesis, which is a biological process. Therefore, autotrophic (124) non-photosynthetic an-aerobic prokaryotes are the only forms of life that could in theory originate independently from anorganic building blocks. They get their energy by chemosynthesis. Chemosynthesis can use simple molecules like hydrogen sulfide, methane (128), hydrogen, ammonium, ferrous iron and manganese(II) as an energy source. This may occur with or without the presence of oxygen. An example is: litho-autotrophic microbes (literal: rock-eating). These bacteria survive on nothing more than oxygen-free hot water, carbon dioxide (CO2) and basalt (84: p.189), (120). In the presence of abiotically derived oxygen, chemolithoautotrophic microorganisms could perform CO2 fixation into cellular biomass dependent on manganese oxidation (129). (Manganese is one of the most abundant elements on Earth). These could be a candidate for independent origin. However, the very first lifeforms could not contain complex DNA and proteins.
- > In fact, energy requirements alone rule out the spontaneous origin of all present-day species. The spontaneous origin of lifeforms much simpler than bacteria is quite another matter (97). However, both Senapathy and Schwabe try to explain present-day organisms exclusively and are unable to consider lifeforms that are significantly simpler than any known lifeforms. Sofar, these results renders any theory of independent origin of species for 99.99% impotent. Furthermore, considering the amazing fact that Senapathy and Schwabe failed to analyse, let alone solve, the real problems inherent in the origin of life, there is nothing left of any value in their theories.
— — —
Tyring to view the biological world through the eyes of independent origin revealed in stead a world in which
organisms depend on previous generations (development, parental care) and other species (ecology). It is amazing and instructive to see how a wide variety of facts, seemingly unrelated to the origins question, such as the boot problem and maternal effects, unexpectedly turn into evidence against independent origin. The biological facts themselves point forcefully to the necessity of an uninterrupted chain of generations. The listed facts are not a complete proof of common descent of all life (77), but certainly are a nearly complete disproof of any form of independent origin, including special creation. These facts make multiple independent origin technically impossible.
To side-step an evolutionary bias, I only used the properties of life as we know it. Therefore, I did not dismiss independent origin by invoking the truth of evolution. Working with an alternative (but wrong) scientific worldview is rewarding and fun.
It produces new insights into the nature of life. Now we have eliminated the impossible. Now we are ready to study the possible origin of life.
References
- Daniel Howard; Stewart Berlocher (1998) Endless Forms.
- Sarah Blaffer Hrdy (1999) Mother Nature. Maternal instincts and how they shape the human species
- Richard Musser et al (2002) "Herbivory: caterplillar saliva beats plant defences", Nature.
- Mike Speed et al (2002) "Animal behavior: Evolution of suicidal signals", Nature. [conspicuous colorations in toxic spiky insects, venomous snakes and frogs]
- Judson et al (2002): "A global disorder of imprinting in the human female germ line", Nature.
- Arie van Noordwijk (2002): Evolutionary Biology: The tale of the parasitic cuckoos. Nature.
- Russell F. Doolittle (2002) Biodiversity; Microbial genomes mulitply. Nature.
- Laurence Hurst (2001) Evolutionary genomics: Sex and the X. Nature.
- Sarah P. Otto, Philippe Jarne (2001) Haploids--Hapless or Happening?" Science. However, the haploid state of the spider mite is a derived state.
- Michael S Brainard and Allison J Doupe: "What songbirds teach us about learning", Nature.
- E. O. Wilson (2000) Sociobiology, chapter 16: Parental Care. pp.336-352.
- Alaska King Crabs website.
- http://www.ecy.wa.gov/programs/sea/pugetsound/species/eelgrass.html
- Barry Sinervo (1998) The Evolution of Male Parental Care in Sticklebacks, Pipefish, and Seahorses (website).
- Mark Ridley (2000) Mendels Demon, Weidenfeld & Nicolson, pp.142-166. (mandatory reading)
- Richard Mann and Fernando Casares (2002) Developmental Biology: Signalling legacies, Nature.
- Jonathan Knight: 'Artifical wombs: An out of body experience', Nature.
- Helena Cronin (1994) The Ant and the Peacock, p.294
- http://www.marinelab.sarasota.fl.us/OCTOPI.HTM
- Henry Gee (2002) "Vegetal vampires stalk South America", Nature.
- Jennie Dusheck (2002) Evolutionary developmetal biology: It's the ecology, stupid!, Nature.
- Stephen Budiansky (2002) "Creatures of Our Own Making", Science.
- David Brickford (2002) "Animal behavior: Male parenting of New Guinea froglets", Nature.
- Dyann F. Wirth (2002) The parasite genome: biological revelations, Nature.
- Ernst Mayr (1988) Towards a new philosophy of biology, p.406.
- http://www.time.com/time/health/article/0,8599,386873,00.html
- "Mice born of lab-raised eggs". Nature Science Update Aug 1 2002.
- Scott Gilbert (2000) Developmental Biology, sixth edition, p.126.
- 'The Role of Maternal Effect Genes in Axis Formation' (no longer available), and The Maternal Effect Genes (9th edition).
- John Maynard Smith & Eörs Szathmáry (1999) The Origins of Life, p. 55. Although modern photosynthesis is one of the most complicated molecular machines we know, there are simpler (and less efficient) forms of light-harvesting, see Plaxco & Gross (2006) Astrobiology: A Brief Introduction, p.154. Most researchers accept that nonoxygenic photosynthesis (which is simpler) arose first, probably shortly after life originated more than 3.8 billion years ago. Life needs an energy source, and the sun is the only ubiquitous and reliable energy source. Nisbet concludes that oxygen-making photosynthesis began at least 2.9 billion years ago. Science, 6 March, 2009 (On the Origin of Photosynthesis, Mitch Leslie).
- See: 'Christiane Nüsslein-Volhard and Drosophila Embryogenesis' in chapter 6 of Developmental Biology, ninth edition. See also her popular science book Coming to Life: How Genes Drive Development (2006). The bicoid gene of Drosophila melanogaster is involved in a variety of early developmental and biochemical processes. Many studies have examined its activity as a morphogen. bicoid mRNA is maternally deposited into the egg and transported to the anterior side, forming a protein gradient that activates transcription of genes in a concentration-dependent manner. Bicoid-deficient embryos cannot develop a head or thorax.
- Paria et al (2002) Deciphering the cross-talk of implantation: advances and challenges. Science.
- Jack Schultz (2002) Biochemical ecology: how plants fight dirty. Nature.
- Patrick Keeling (2001) Parasites go the full monty. Nature.
- Joshua Tewksburt (2001) Seed dispersal: Directed deterrence by capsaicin in chillies. Nature.
- J. Allan Downie (2001) Genome sequencing: The ABC of symbiosis. Nature.
- Evelyn Strauss (2007) 'Detoxifying Enzyme Helps Animals Stomach Bacteria', Science, 21 Dec 2007.
- see note 32.
- Pierre de Wit (2002) Plant biology: On guard. Nature.
- M Azim Surani (1999) Imprinting (Mammals) Encyclopedia of Life Sciences.
- see: vicious circle.
- A. Fazleabas (2003) What makes an Embryo Stick? Science.
- Elizabeth Pennisi (2003) On Ant Farm, a Threesome Coevolves, Science.
- C.R. Townsend, M. Begon, J.L. Harper (2003) Essentials of Ecology, second edition, p224.
- N.B. Davies (2000) Cuckoos, cowbirds and other cheats, p.25.
- Wallace Arther (2000) The Origin of Animal Body Plans (review).
- Carina Dennis and Richard Gallagher (2001) The Human Genome, Nature Publishing Group, hardb. 140 pp.
- Jean Marx (2002) Telomeres: Chromosome End Game Draws a Crowd. Science.
- John Maynard Smith & Eörs Szathmáry (1999) The Origins of Life, pp.91-93.
- Jian Xu et al (2003) "A Genomic View of the Human-Bacteroides thetaiotaomicron Symbiosis", Science. See also: Steven R. Gill et all (2006) "Metagenomic Analysis of the Human Distal Gut Microbiome", Science.
- A review of E. O. Wilson's "Pheidole in the New World. A Dominant, Hyperdiverse Ant Genus", Science.
- Jared Diamond (1999) Guns, Germs, and Steel, W.W. Norton paperback, page 116.
- Lynn Helena Caporale (2003) Darwin in the Genome. Molecular strategies in biological evolution, McGraw-Hill, p.48.
- Wallace Arthur (2000) The Origin of Animal Body Plans, p. 120-121.
- Shirley M. Tilghman (1999) The Sins of the Fathers and Mothers: Genomic Imprinting in Mammalian Development, Cell, Vol. 96, 185-193, January 22, 1999.
- Carl Zimmer (2001) Evolution. The Triumph of an Idea, Chapter 8: Coevolution, pp.189-209. This chapter is obligatory reading.
- Jared Diamond (1987) "Aristotle's theory of mammalian teat number is confirmed.", Nature.
- Edward O. Wilson (2001) The Diversity of Life, new edition, p.217.
- Lynn Margulis (1998) Five Kingdoms, p.364.
- There is a very interesting and relevant article "The facts of Life", NewScientist (2003) about minimal genomes and how to create life in the testtube starting with pure chemicals.
- Graur and Li (2000) Fundamentals of Molecular Evolution, p.158.
- David Bainbridge (2003) The X in sex. How the X-chromosome controls our lives.
- Mark T. Ross et al (2005) "The DNA sequence of the human X chromosome", Nature, and Bruce Lahn (1999) "Four evolutionary strata on the human X chromosome", Science.
- W. Wayt Gibbs (2003) "Untangling the roots of cancer", Scientific American, June 2003.
- David Haig (2002) Genomic imprinting and kinship, Rutgers University Press, p.23. See also: Tomohiro Kono et al (2004) Birth of parthenogenetic mice that can develop to adulthood: "Our study shows that imprinting is a barrier to parthenogenetic development in mice". See also: David A. F. Loebel and Patrick P. L. Tam (2006) Mice without a father: "In mammals, genomes from both parents are generally needed to make viable offspring. But changing the expression of 'imprinted' genes can render the father's contribution dispensable".
- Jessica Downs (2003) "Cancer: Protective packaging for DNA", Nature.
- The laws that Mendel discovered are ultimately based on the diploid state of the organisms he studied. Some historians claim that Mendel did not fully understood this (see: John Waller)
- John Allman (2000) Evolving Brains, Scientific American Library, paperback, 224 pages (page 175).
- Christopher Wills and Jeffrey Bada (2000) The Spark of Life. Darwin and the Primeval Soup, p. 196.
- Russell Foster & Leon Kreitzman (2004) Rythms of Life: The Biological Clocks that Control Daily Lives of Every Living Thing: a popular science book reviewed in Nature. See also: Circadian Rhythms (introduction webpage). Genetic and biochemical screens identify the same protein, which determines period length of the Mammalian circadian clock by degradation of a known component (Science 11 May 2007). The circadian clock in fruit flies is encoded by a set of transcriptional activators and repressors that comprise an autoregulatory transcription-translation feedback loop.
- John Maynard Smith and Eörs Szathmáry (1999) The Origins of Life, page 55. This book continues to deliver new insights, that is fundamental insights into what life is.
- Christian de Duve (2002) Evolving Life, p. 130.
- Carina Dennis (2003) Nature.
- For a good discussion of the basic principles of energy metabolism and their application to the origin of life, see: Peter Skelton (1993,1996) Evolution. A Biological and Palaeontological approach, page 870-881.
- Nir Ohad (2007) 'Plant development: Parental conflict overcome', Nature 447, 275-276 (17 May 2007). Matters are still more complicated than the above description.
- David Bainbridge (2000) Making babies. The science of pregnancy, pp 45-51.
- See why anything less than full common descent of all life is contradicting facts and logic Chapter 3 of Why Intelligent Design Fails (review).
- Observations show, that impairment of normal parental imprinting may interfere with cellular differentiation and thereby increase the probability of cancerous development (colon cancer). George Klein, Nature.
- Laura Carrel & Huntington Willard (2005) Nature.
- Eva Jablonka and Marion Lamb (2005) Evolution in Four Dimensions, p.168. (review).
- It is impossible to start a species with an independently born male and female, because it is highly unlikely that a second individual of the opposite sex originated independently with a genome similar enough to mate successfully with the first individual and additionally, have fertile offspring. In the Bible Eve was designed and created specifically to match Adam, but this trick cannot be used in a scientific theory.
- Wolfgang Driever (2005) A Message to the back side Nature. Inhibiting the synthesis of protein from the maternal as well as the zygote ndr1 mRNA produces radially symmetric embryos!
- Garry Hamilton (2006) "The Gene Weavers", Nature
- Kevin W. Plaxco & Michael Gross (2006) Astrobiology : A Brief Introduction.
- Christiane Nüsslein-Volhard (2006) Coming to Life: How Genes Drive Development.
- Geerat J. Vermeij (2004) Nature. An economic history, p.87.
- Science, 29 October 1999, p. 877.
- Luis M. Márquez et al (2007) 'A Virus in a Fungus in a Plant: Three-Way Symbiosis Required for Thermal Tolerance', Science 26 January 2007: 513-515.
- Debra Lieberman, John Tooby and Leda Cosmides (2007) The architecture of human kin detection, Nature 445, 727-731 (15 February 2007)
- Nancy A. Moran (2007) 'Symbiosis as an adaptive process and source of phenotypic complexity', PNAS, published online May 9, 2007. (free access)
- Laurie E. Comstock (2007) 'Microbiology: The inside story', Nature 2 Aug 2007.
- Andreas Brune (2007) 'Microbiology: Woodworker's digest', Nature 22 Nov 2007.
- John Odling-Smee, Kevin Laland, Marcus Feldman (2003) Niche construction. The neglected process in evolution, page 10.
- Sugako Ogushi (2008) 'The Maternal Nucleolus Is Essential for Early Embryonic Development in Mammals', Science 1 February 2008.
- Vaclav Smil (2008) Energy in Nature and Society. General energetics of complex systems, p.123, 113, 112.
- Elizabeth Pennisi (2008) 'Bacteria Are Picky About Their Homes on Human Skin', Science, 23 May 2008.
- Famous origin-of-life reseacher David W. Deamer pointed out that much simpler membranous vesicles can can be permeable enough to let a cell grow by allowing small molecules through, yet stable enough to maintain the integrity of the cell. See: David W. Deamer (2008) 'Origins of life: How leaky were primitive cells?', Nature, 3 July 2008.
- Julius Brennecke et al (2008) 'An Epigenetic Role for Maternally Inherited piRNAs in Transposon Silencing', Science 28 November 2008.
- Michael Ruse, Joseph Traivs (2009) Evolution. The First Four Billion Years, page 850.
- The complex relationship between the "Holstein" butterfly (Heliconius sapho), its food plant and its pollen source (accessed 16 Feb 2009)
- Ueli Grossniklaus (2009) Paternal Patterning Cue, Science 13 March 2009
- Donald Kroodsma (2007) The Singing Life of Birds, Houghton Mifflin Company, paperback. p.50.
- Alessandro Minelli (2009) Forms of Becoming. The evolutionary biology of development p.202-203.
- Nipam H. Patel (2009) 'Developmental biology: Asymmetry with a twist', Nature.
- Justin L. Sonnenburg (2010) 'Microbiology: Genetic pot luck', Nature, 8 April 2010
- Hanna Kokko, Michael Jennions (2010) Behavioural ecology: Ways to raise tadpoles, Nature, 15 Apr 2010.
- K. S. Hayes et al (2010) 'Exploitation of the Intestinal Microflora by the Parasitic Nematode
Trichuris muris', Science 11 Jun 2010.
- Frequently Asked Questions about Animal Rehabilitation and Release.
- Liping Zhao (2010) 'Genomics: The tale of our other genome', Nature, 465, 879-880 (17 June 2010)
- Other researchers claim that babies born via cesarean section, experience a more sterile entry into the world than those born vaginally.
- Conservation Biology: Pandas prefer their trees mature, Nature, 469, 268 (20 January 2011)
- Elizabeth Pennisi (2011) Going Viral: Exploring the Role Of Viruses in Our Bodies, Science 25 Mar 2011
- F.M. Jiggins, G.D.D. Hurst (2011) Rapid Insect Evolution by Symbiont Transfer, Science 8 April 2011
- Arbel D. Tadmor et al (2011) Probing Individual Environmental Bacteria for Viruses by Using Microfluidic Digital PCR, Science 1 Jul 2011
- Marc-André Selosse, François Rousset (2011) The Plant-Fungal Marketplace, Science 12 Aug 2011.
- John P. McCutcheon, Carol D. von Dohlen (2011) An Interdependent Metabolic Patchwork in the Nested Symbiosis of Mealybugs. Current Biology August 11, 2011
- Anna Ajduk (2011) Rhythmic actomyosin-driven contractions induced by sperm entry predict mammalian embryo viability, Nature Communications 2 09 August 2011
- Sperm Spills Its RNA Secret, Science 9 September 2002
- Arthur L. Beaudet (2012) Angelman syndrome: Drugs to awaken a paternal gene, Nature 481, 150–152 (12 January 2012)
- Interestingly, those cells succeed in producing biomass (carbohydrates) by using CO2 reduction just like photosynthesis in all green plants, but then of course without photosynthesis.
- Robert M. Brucker, Seth R. Bordenstein (2013) The Hologenomic Basis of Speciation: Gut Bacteria Cause Hybrid Lethality in the Genus Nasonia, Science 9 August 2013. This is a special anti-independent origin case: additionally, Nasonia offspring are oviposited by the wasp mother inside the puparium of the fly, where the eggs develop to adulthood before emerging from the fly host in about 14 days. So, the wasp is parasitic (wiki).
- Cryptomonas Microbe Wiki.
- Timothy L Karr Centrosome inheritance: A central 'in-egg-ma' solved? Current Biology, Volume 11, Issue 1, 9 January 2001
- "The earliest ancestors [of life] were autotrophs, for heterotrophy imposes demands on the chemical composition of the environment for specificity and concentration that cannot be fulfilled by any known geochemical processes." Harold Morowitz (1999) 'A Theory of Biochemical Organization, Metabolic Pathways, and Evolution', Complexity pp 39-53. 7 Jul 2014
- Derk-Jan Dijk, Anne C. Skeldon (2015) Biological rhythms: Human sleep before the industrial era, Nature 12 Nov 2015
- However, little evolutionary effort would be required to evolve anoxygeneic photosynthesis, and it could have started soon after the origin of life itself. Karo Michaelian (2016) Thermodynamic Dissipation Theory of the Origin and Evolution of Life, p.84.
- Cassandra G. Extavour, Michael Akam (2003) Mechanisms of germ cell specification across the metazoans: epigenesis and preformation.
- Bacterium Methylococcus capsulatus is a prokaryote that metabolize methane CH4 as their only source of carbon and energy.
- Hang Yu, Jared R. Leadbetter (2020) Bacterial chemolithoautotrophy via manganese oxidation, Nature 583, pages 453-458(2020). 16 Jul 2020
- Purple Earth hypothesis. 18 Oct 2020
Recommended reading
If I have to single out any books for recommended reading, these 4 are certainly included:
- Chapter 7: 'Organisms as Habitats' of Townsend (2003) Essentials of Ecology if you think organisms are independent individuals. Alternatively: note 100.
- Sarah Blaffer Hrdy (1999) Mother Nature for anybody who thinks independent origin (of humans or mammals) is possible (see short review)
- Chapter 8: 'Co-evolution', of Carl Zimmer (2001) Evolution. The Triumph of an Idea in order to see that organisms are adapted to each other, which is a priori a barrier to independent origin.
- Lynn Margulis and Dorion Sagan (2002) Acquiring Genomes. A theory of the origin of species. If anything contradicts independent origin then it is Margulis' now well established symbiosis theory (review).
- The nature of the last universal common ancestor and its impact on the early Earth system, Nature 12 July 2024. Very important publication. The Last Universal Common Ancestor (LUCA) very likely had a genome of 2.75 Mb and 2600 proteins and lived 4,2 billion years ago. The earth formed 4,5 billion years ago. So there was only 300 million years to evolve LUCA from abiotic molecules. 12 July 2024
Further Reading
|


{kind=link}