Was
Darwin
Wrong?
Was Darwin Wrong? |
Home | Intro | About | Feedback | Prev | Next |
|
A Chemist's View of Life: Ultimate Reductionism & Dissent
A review of Christian Schwabe's 'The Genomic Potential Hypothesis' by Gert Korthof. Embryos swimming in a fresh water pond?
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
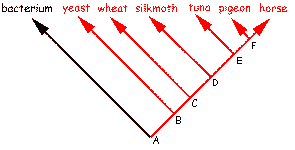
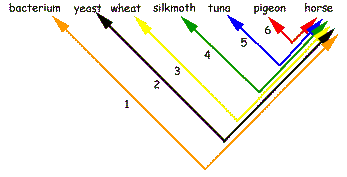
Two basic interpretations of the relations between species |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
The difference between Darwinism and Schwabe could not be more fundamental than it is. Independent Origin can easily explain dissimilarities in species. Diversity is expected when every species arose directly from non-life. Schwabe chooses Independent Origin of all species because he sees irreparable difficulties with particular phylogenetic trees of genes/proteins. This choice contradicts the principle of Common Descent of all life. The tree of life simply does not exist. Schwabe has a number of additional arguments pro Independent Origin (see table). IO needs to explain similarities in organisms too. If all species arose independently then why do similarities exist at all? Why groups within groups? Why are humans within a group of apes within a group of mammals within a group of vertebrates within a group of animals within a group of eukaryotes? Sequence similarities must be accidental in the Independent Origin theory (1). According to Darwinism there are too much similarities in species to be explained by chance. Although Schwabe does not state it explicitly, his view implies for example that humans and chimps (>95% genetic similarity) arose independently. All birds arose independently. All birds of prey arose independently. All species of eagle arose independently. All this is absurd in the eyes of Darwinists. Furthermore if all species arose independently, why do they all have the same genetic code? A multitude of alternative codes is compatible with life [see: review of Remine's book]. Schwabe's view can best be summarized by comparing it with Darwinism:
All topics in the table are logically connected. Whatever the facts, the logic of the paradigm demands some statements to be true and others to be false. For example it's logical in the Independent Origin paradigm to accept fixity of species because species don't need to change. They don't need to split, because all species originated simultaneously. But then natural selection isn't necessary anymore: species are not created by natural selection. Mutations aren't necessary either. Anyway mutations cannot reshape one species into another.
Since mutations are the source of variation, variations must be meaningless. So variation cannot be not created during the course of evolution, it must be present right from the origin of life. And if mutation, natural selection, and variation are unimportant in evolution, they cannot explain adaptation. This is remarkable, because any big theory about life has to be able to explain adaptation (29). Independent Origin does not need intermediates, because species are not produced by a gradual process of small modifications. There is no problem of missing links, because Independent Origin does not need links. Since the Independent Origin paradigm accepts the fossil record as it is, it accepts extinction. All species go extinct and no new species will ever appear in the course of evolution. When new species seem to appear in the fossil record, they cannot be really new, but they arise from pre-existing 'pro-forms'. It follows from the Independent Origin paradigm that the particular genetic code on earth must be natural, reproducible and truly universal. It also follows from I.O. that all DNA and protein sequences arose independently and not from duplications + mutations. Although it is not a necessity, it's no surprise that chemistry has the power to create the millions of species independently over and over again and that all properties of species are directly explained by chemistry, since Schwabe is a chemist. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Origin of life |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Schwabe's ideas about the origin of species differ radically from those of mainstream scientists. Mainstream science postulates one origin of life (almost by necessity). Schwabe claims that all life arose directly from pools of chemicals in a natural way. He points out that the Miller-Urey experiment produced amino acids in exactly the same proportions as found in the Murchison meteorite. This means that truly universal laws of chemistry are at work. Those laws are favourable for the origin of life. More than favourable: those laws make the origin of life inevitable when the proper reaction conditions prevail. Schwabe wants to make the origin of life research "a hard science based upon chemistry, thermodynamics, kinetics and the laws of mass action". The origin of life is not a one-time lucky event "in contrast to the chance-oriented Darwinian paradigm". Only after reading Gánti (23) I realized that Schwabe did not analyse the task at hand: what is it that needs to be explained? What is life? And if you skip that question, how can you make a significant contribution to the origin of life field? Moreover, how can you ever hope to explain the origin of millions of complex species, if you do not have a clear understanding of how to explain simple life forms? There is more about the problems and dilemmas of the origin of life in books for the non-specialist, for example Fry (26), than in Schwabe's book! Schwabe is blind to the fact that he and scientists in general have a common task: the search for natural explanations of life. Schwabe prefers an unproductive ideological battle against Darwinists above producing 'hard science'. Furthermore, his ideological points are not new, sometimes trivial and not rejected by everybody. For example Nobel Prize winner Christian de Duve already stated that life did not arise by chance, and chemistry is deterministic. Schwabe seems to be unaware of the fact that chance does not exclude inevitability. For example there is a 99.9% probability that number six will occur in a series of 38 throws of dice (2). Even in the evolution of species on earth there is inevitability despite a lot of historical contingency according to palaeontologist Simon Conway Morris. This is because of convergence, and convergence means independent origin of similar characteristics. Many others before Schwabe have pointed out the lack of laws in biology. For example in the morphologies of organisms (Brian Goodwin) and the origin of life (Stuart Kauffman). So Schwabe's campaign against chance is not very original. Finally, Schwabe's non-evolutionary theory of the origin of life has deep roots in Greek philosophy. Philosopher and physician Hippocrates, unable to think in terms of evolution, "did not search for the possible mode of the origin of the simplest organisms on earth, but sought to derive man himself, the crown of earthly existence, directly from material substances"! (22). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Origin of the genetic code |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
The 'genetic code' is the translation of the 64 base triplets into 20 amino acids. Both Schwabe and mainstream science accept that the genetic code is universal for life. However the explanation of this fact could not be more different. For mainstream science the genetic code was a 'frozen accident', because it did not seem to change and it seemed to be an arbitrary code.
What if Schwabe were right about the inevitability of the code? A proof of the inevitability of the code would not destroy common descent, but it would stop to be evidence for common descent. However, the failure to prove the inevitability of the code would make independent origin very implausible. What about the variations of the code? Schwabe's position and Darwinism are in fact extreme positions and I think there is a middle position: the genetic code is neither completely inevitable, nor completely arbitrary. The genetic code couldn't be 100% inevitable anyway because there are a number of small variations of the standard code (24). Schwabe ignores this. The existence of small variations of the code imply that the code is neither completely frozen and nor completely universal. However, "the existence of non-standard codes offers no support for a 'multiple origins' view of life on earth. Lineages that exhibit non-standard codes are clearly and unambiguously related to organisms that use the standard code: they are distributed as small 'twigs' within the evolutionary tree of life" (28). Moreover, the small variations suggest evolvability of the code. There is computational evidence that the code has been optimised for error minimisation by natural selection (25). Schwabe's theory of the origin of the genetic code has a definite disadvantage when compared to an evolutionary theory. Schwabe cannot start with a simple code and evolve a complex one. Schwabe's theory must start with all 20 amino acids, because all species now have 20 amino acids in the code. However, an evolutionary scenario can start with a limited number of amino acids (6-10) and gradually increase the number to 20 amino acids (30). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
The Genomic Potential Hypothesis |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
For Schwabe the origin of life and the origin of species are the same problem and he tries to eliminate the 'lucky accident' model from the origin of life as well from the origin of species. I have sympathy for his rejection of 'lucky accidents' in the origin of life.
In the domain of the origin of life obviously the main role is played by chemistry. If this attitude helps to explain the origin of the genetic code, it would be a tremendous scientific progress. Successful or not, it is good science to try to do so. However his attempt to extend this approach to the origin of species fails, because the 'lucky accidents' reappear in full magnitude in his theory of the origin of genomes, as I will explain below. How can chemistry of nucleotides explain the origin of genomes? It is quite a different matter to explain the origin of life by chemistry than to explain the origin of fully functional genomes by pure chemistry (23). Let's first see his views about the origin of genomes:So in Schwabe's model of the origin of life large numbers of bases + relative concentration of G,C,A,T + chemical laws + suitable conditions are enough to create not only the simplest but all the genomes. Schwabe seems to think that if a small simple genome (a single cell) can be generated than a large complex genome (of a multicellular organism) can be generated too. But the laws of chemistry cannot explain a DNA sequence. This is a crucial point. Paul Davies most forcefully and clearly explained this crucial point (3,p120). Davies conclusion is that 'Specific randomness' [DNA] cannot be produced by a deterministic mechanical lawlike process without a huge element of luck (i.e. chance) (3,p. 119). Indeed, physics and chemistry can explain the structure and dynamics of DNA bases; why DNA is a stable double helix, why DNA is so perfectly able to replicate itself. However chemistry is powerless to determine the subset of all possible DNA sequences that are biologically significant (4). Only a tiny fraction of all random DNA sequences have biological meaning. So if Schwabe wants to get rid of luck, he needs a mechanism other than the laws of chemistry. Schwabe has difficult times explaining genomes without natural selection. He comes in conflict with his own probability calculations on the origin of relaxin from insulin by gradual steps. To convert both insulin (50 amino acids) into relaxin and the insulin receptor (1000 amino acids) into a relaxin receptor, one needs 4x101050 trials in total. Such probabilities mean that such an event would not happen according to Schwabe. But if it is impossible to evolve one hormone/receptor pair by modification of existing ones, why should it be easier to create all the thousands of genes of an organism in one independent origin event? This needs to be 1 trial otherwise natural selection would be implied. If it is difficult for Darwinian organisms to 'find' those sequences, why would it be more probable in a pool of nucleotides? I can think of only one advantage: a litre water can contain more random DNA chains than a population of organisms (a species). Organisms are subjected to more restrictions than molecules. Organisms need space, food, time to reproduce and grow. A million pools of 1 litre water can contain more DNA sequences than a million species. And species are slower than molecules. But both species and naked DNA sequences live in the same abstract 'DNA space' and the same physical world. The number of all possible sequences of say 1 billion bases long (a genome length) is the same in both theories. And so is the fraction that is viable. Anyhow, the assembled genome needs to be tested for its viability in both theories. To test a potential genome one would have it sitting in a cell and see if it survives in its environment. This is the only way for single celled organisms. But how to test multicellular organisms in a single cell state as Schwabe wants it in his Genomic Potential Hypothesis? Impossible. There is no short cut for testing genomes. I have the idea that Schwabe is trying to substitute extinction for natural selection as the mechanism of eliminating wrong genomes. If so, he should admit this. The worst thing is that Schwabe, as an exact scientist (chemistry being more exact than biology!), does not even attempt to create a rigorous foundation for his alternative theory. I have only touched upon one of the massive amount of objections to The Genomic Potential Hypothesis. I add just a second reason in the next paragraph 'Reductionism'. There are more examples in my review of Senapathy, who has a nearly identical theory called 'Independent Birth of Organisms'. Furthermore, many fundamental facts about how organisms are constructed clash with the whole idea of independent origin of genomes (here is a summary). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Reductionism: where are the chromosomes? |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
In Schwabe's philosophy organisms are reduced to genomes and genomes are reduced to DNA-sequences. Here is reductionism in a nutshell: "Once they [nucleotides] are linked into chains they spell out an organism". But a genome is not just a very long DNA sequence.
It would be foolish to say that karyotypes arose simultaneously and independently from a 'pool of fresh water' and produces the similarities and dissimilarities we observe now. It would be still more foolish to claim that for example the (human) sex chromosomes X and Y arose spontaneously from 'pools of fresh water'. The function of the highly differentiated sex chromosomes and the specific genes they contain can only be understood from their evolutionary history. Evidence supports the theory that the XY pair of chromosomes evolved from a non-differentiated pair of chromosomes, and that the Y chromosome degenerated to a minimalist endproduct but is still essential for creating a male (19). There is a large body of literature on the evolution of sex chromosomes and the fundamental biological principles underlying its peculiarities. Not a topic to ignore if you want to explain genomes. But still this is only a minor part of the literature about chromosomes in the animal and plant kingdom. Each chromosome is evidence against independent origin of genomes. I could write a whole book on this topic. And then the omission of chromosomes is only one example of Schwabe's reductionism. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Pro-Forms: hypothetical or imaginary? |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
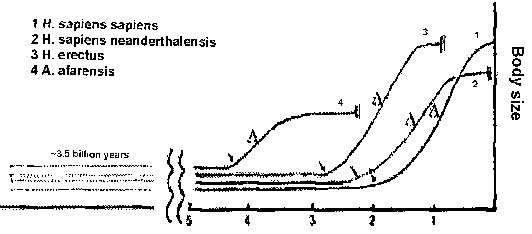
In Schwabe's theory every species originated 3.5 billion years ago. This should show up in the fossil record. But it does not.
So Schwabe invented the concept 'Pro-Forms' ('without form', 'before form'), a concept unknown in biology. The concept is crucial in his theory, but there is no single chapter devoted to 'pro-forms'. Information about 'pro-forms' is vague and hidden in remarks, metaphors and graphs. Schwabe views the relation between 'pro-forms' and adult animals as the relation between a caterpillar and a butterfly. Just as a single insect genome produces a caterpillar and a butterfly, a single animal genome produces 'pro-forms' and an adult animal. But since Schwabe claims that every species started 3.5 billion years ago, they must exist for millions and billions of years in a formless (single cell?) state. After that period of invisibility, adult animals become visible in the fossil record. This immediately causes a problem: maintaining genomes for 3.5 billion years in good order. Only natural selection can prevent an accumulation of mutations in genes, and genes can only be selected if they are used ('use it or lose it' principle). Genes involved in an adult multicellular individual are not used in a single cell state (principle of cell differentiation).
What does that mean for humans? Schwabe accepts the standard age of the human species (about 5 million years). For Schwabe this means humans became visible at that time, since humans existed 3.5 billion years as very small 'pro-forms' and started to increase in size during the last 2 million years. According to a graph (fig. 5 in Chapter 12), this increase in body size is gradual from invisible to our current size. This implies that humans lived as fly-sized, rat-sized, cat-sized, dog-sized creatures! Dwarfs? Midgets? Gnomes? Does Schwabe really look for adult rat-sized human skeletons in the fossil record? Are there male and female 'pro-forms'? Are they haploid or diploid? How do they reproduce? There are many steps in Professor Schwabe's contention which are not stated with as much clarity and precision as one could desire, and I fear the reason is that clarity and precision make them implausible. The 'pro-form' idea seems merely an ad hoc explanation to protect the theory of independent origin from falsification by the data of the fossil record and has no positive data that support it. There is another problem for Schwabe: oxygen. Either human cells originated 3.5 billion years ago and were anaerobic (don't need oxygen), or they originated only after oxygen levels reached current levels of 21% (about 550 million years ago) and were aerobic (need oxygen). Whether humans existed as single cells or multicellular beings, in both cases human cells need oxygen. Human cells are aerobic. That is the way human cells are. Therefore, the cells in the first scenario are not human cells. The second scenario contradicts Schwabe's assertion that all genomes originated 3.5 billion years ago. If Schwabe insists on 3.5 billion years, then he is forced to accept an evolutionary change from anaerobic to aerobic state. This conflicts with his non-evolutionary worldview, because in his worldview genomes are essentially fixed after they come into being. Even if a genome was accidentally pre-adapted to a future oxygen-rich atmosphere, it simple could not survive until then (31). A courageous dissident would have mentioned all this. Let's assume it escaped his attention. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Embryos swimming in a fresh water pond |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
The most catastrophic implication of his theory of independent origin of genomes is that a human fetus must develop outside the womb in a fresh water pond. There is no way around. If your theory demands that a mammalian embryo develops from a single cell and that cell originates directly from a fresh water pond, then this implies that somehow, somewhere, sometime a transition must take place from an embryo living in a water-based environment and to an air breathing baby. Further implications are that there is no maternal womb support, so no constant body temperature, no constant food and oxygen supply, no mother's milk supply after 'birth' (what does 'birth' mean in this context?), no parental care. Nothing! Nine months! Internal gestation in all placental mammals invented for nothing. Dispensable! In Darwin's time creationists wrestled with the explanation of belly buttons of the first humans (20). Is Schwabe unaware of all this? One gets cynical when confronted with all the absurd consequences of independent origin. Is this science fiction? Is this a chemist with too much imagination? Birds and even some fish species have parental care. Even reptiles do something to ensure egg survival. Even plants invest food in their seeds, etc. I collected all these fundamental and fatal objections on the page Independent Origin and the facts of biology. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Where did the first cells got their energy from? |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
The most straightforward and elegant refutation of independent origin of all species is the energy requirement of the first life forms.
Cells get their energy in one of three ways (27):
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Schwabe's problems with Molecular Evolution |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Why invent a scientific alternative for evolution? Consider the following two DNA sequences of 10 bases:
Let's have a look at his criticism of Darwinism. Schwabe is a chemist and his expertise is the hormone relaxin. He published a standard work on relaxin (6). He studied relaxin sequences and their biochemical activities in different animals. He claims that distribution patterns of insulin, relaxin and cytochrome in different species contradict common descent. For example
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
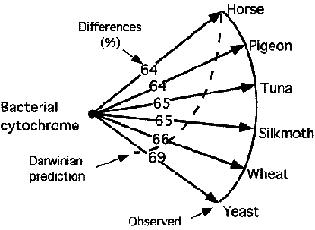
The Schwabe-Denton interpretation of cytochrome-c differences |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Remarkably Schwabe repeats Michael Denton(1985)'s illustration of cytochrome differences (without acknowledging the source of his illustration) and he repeats the same interpretation errors as Denton (21).
The errors are:
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Intermediates: whose problem? |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Schwabe states that Darwinism necessarily predicts intermediates in the fossil record as well as in terms of protein structures, but the intermediates remained elusive. Indeed Darwinism expected intermediates. Nature does not make jumps. Jumps would be miracles. Do jumps in anatomy and morphology exist? This is an empirical and experimental question. The difference of a six-legged fruitfly and a many legged velvet worm (two distantly related groups) seems a big jump. However, as recently has been found small mutations can turn a many legged creature into a six-legged creature (12). So there never can be an intermediate between six-legged and a xx-legged form. So it's pointless to search for such an intermediate. Although it is not demonstrated that no intermediates in general are necessary in evolution, the Darwinian expectations about intermediate forms have to be re-adjusted. This is for example clear from enzyme functions. Major shifts in enzyme function may often require no more than a few changes: To change lactate dehydrogenase into a malate dehydrogenase takes only one amino acid substitution (American Scientist 1998 Jan-Feb). "There is evidence that proteins with de novo functions are more likely to be generated from genetically modified proteins rather than from sequences formed at random (Doolittle, 1981, 1987a)" (13). If Schwabe tried to present a balanced review of the status of neo-Darwinism then he should have mentioned these examples. While thinking about these matters it suddenly occurred to me that Schwabe has given no good reasons for the fact that there should be no abundant number of intermediates according to his own model. If genomes are assembled in a random way from pools of nucleotides, then we should expect every possible sequence of DNA, with no gaps. It would be a miracle if only 'the right' genomes originated and nothing more. If we return to the example of the two ten base DNA sequences of the paragraph Schwabe's problems with Molecular Evolution, then we should expect to find 1,048,576 sequences of 10 bases long. More than a million. Where are they? If they are not part of any gene, where are they? This problem is particularly pressing if one indignantly rejects natural selection (as Schwabe does). Furthermore, if genomes are assembled in a random way from pools of nucleotides, how do they manage to produce organisms that can be divided so neatly in distinct Linnean systematic groups? In fact the independent origin model should expect all kind of strange intermediate forms. Birds with scales on their bodies like reptiles or hair like animals, and naked birds, and birds with teeth, mammalian birds, birds with photosynthesis, trees with eyes and brains, etc. We do not find them in nature. Finally, what is more difficult: to turn a reptile into a mammal; a reptile into a bird (evolution) or to produce a mammal or bird from scratch (independent origin)? Obviously, Independent Origin and Evolution need to solve the same fundamentental observation of biology: Adaptation. It is completely absent in Schwabe's theory. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Senapathy's theory: independent? |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Senapathy(14) is only mentioned once in passing by in Chapter 8, but not discussed. This is a remarkable fact and is odd. One would have expected that both authors would have profited from each other's expertise to strengthen their theory (as happens in normal science). We do find nothing of the sort in the books of Schwabe and Senapathy. Schwabe did publish a paper about his genetic potential hypothesis in Perspectives in Biology and Medicine (1984), that is 10 years before Senapathy's book. But I don't know if Senapathy knew this publication. Schwabe is not in the index of Senapathy's book. Is it possible that Senapathy and Schwabe independently invented a nearly identical theory? A wonderful example of Independent Origin! There are a number of differences between the two theories. Senapathy rejects the independent origin of prokaryotes, while accepting it for eukaryotes. I did not find this distinction in Schwabe's theory. Schwabe accepts the independent origin of both prokaryotes and eukaryotes, thereby opposing Senapathy. Another difference is Schwabe's 'pro-forms'. Senapathy does not need them, because species seem to originate during a very long geological period in his theory. Thirdly, Senapathy decided that it was too improbable to accept independent origin of closely related species and accepted common descent for those species. I did not found such a thing in Schwabe. Finally, although both claim that genomes are fixed and do change, Senapathy claims more often that genomes are "immutable", while Schwabe more often holds that genomes 'reorganise' themselves during geological history. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Conclusion |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
In a 114-page book chemist professor Christian Schwabe claims (1) to have produced data that cannot be explained by Darwinism, and (2) to have produced a better explanation for the origin of species (and the origin of life) than Darwinism does. He (3) restricted the origin of all species to a short period 3.5 billion years ago and (4) when confronted with a fossil record which does not show the existence of all species 3.5 billion years ago, he (5) invented 'pro-forms' which remain in a single cell state during million of years and become only visible at precisely the time required by the fossil record. Schwabe neither produces positive evidence for 'Pro-forms', nor solves any of the huge biological and geological problems 'pro-forms' cause. Schwabe's alternative pro-forms theory fails because of technical biological reasons, not because of evolution or Darwinism. However, without those 'pro-forms' his theory is seriously incomplete. His Genomic Potential Hypothesis, is vague, speculative and has nothing to do with the new biological discipline called genomics. The choice between Schwabe's theory and Darwinism is the choice between million independent origins including babies without mothers, the rejection of mutation, natural selection, the tree of life, the reality of speciation, the reality of genera, families, etc on the one hand, and on the other hand a theory that explains adaptation and the diversity of life in great detail, and is in harmony with all biological disciplines, but which has a number of perceived inconsistencies. When a theory clashes with, and is silent about so many basic biological facts, then the choice is easy. The choice is a theory that explains a maximum of data and has a minimum of anomalies and puzzles. Evolution may have its anomalies and puzzles, and is certainly unfinished, but I prefer it above science fiction. As long as the Genomic Potential Hypothesis is in such a deplorable condition, no biologist has the obligation to disprove it. On the contrary, it is Schwabe's obligation to deliver a theory worth disproving. Schwabe is the ultimate dissident. He disagrees with anything and everybody. He doesn't even want to know that he shares a common goal with the origin of life research community (which is distinct from the evolutionary biology community). A common task is that both need to explain the origin of metabolism, membranes, RNA, DNA, the genetic code and how they come together to form life. Why bother at all? Schwabe's hypothesis is the only non-creationist, naturalistic alternative to evolution. In the process of trying to make sense of it, it deepened my insight into life and evolution |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
 The hardback edition (ISBN 1-58706-044-2) is exceptionally expensive (Amazon: $119; BarnesNoble: $144). In 2002 it was available for free online at Eurekah.com, Landes Bioscience. In 2003 I found out that only the abstracts of each chapter are free. Subscription is now necessary. See contents. I used version of 4.18.01.
The hardback edition (ISBN 1-58706-044-2) is exceptionally expensive (Amazon: $119; BarnesNoble: $144). In 2002 it was available for free online at Eurekah.com, Landes Bioscience. In 2003 I found out that only the abstracts of each chapter are free. Subscription is now necessary. See contents. I used version of 4.18.01. This book is my main source for evaluating the Genomic Potential Hypothesis (GPH). The book is somewhat sketchy, not well elaborated. However, this book is the most complete exposition of the GPH. The author should expand the contents, because it's weak at educating the non-specialist reader and he should also educate himself in biological disciplines. In his articles in scientific journals he is more precise and detailed about specific molecular puzzles, but not detailed about his GPH. PostscriptI notified Prof. Schwabe of this review and asked for comments. Up to the present Schwabe did not bother to defend his theory. [ Jan 2003 ]
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||
Notes
Further Reading
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Korthof blogspot | home: wasdarwinwrong.com | wasdarwinwrong.com/korthof56.htm |
| Copyright ©G.Korthof 2002. | First published: 24 Mar 2002 | Updated: 21 Sep 2013 Notes/FR: 2 Jan 2013 |